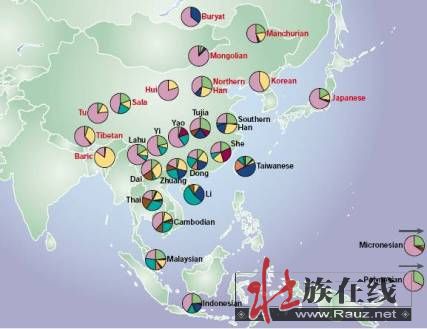
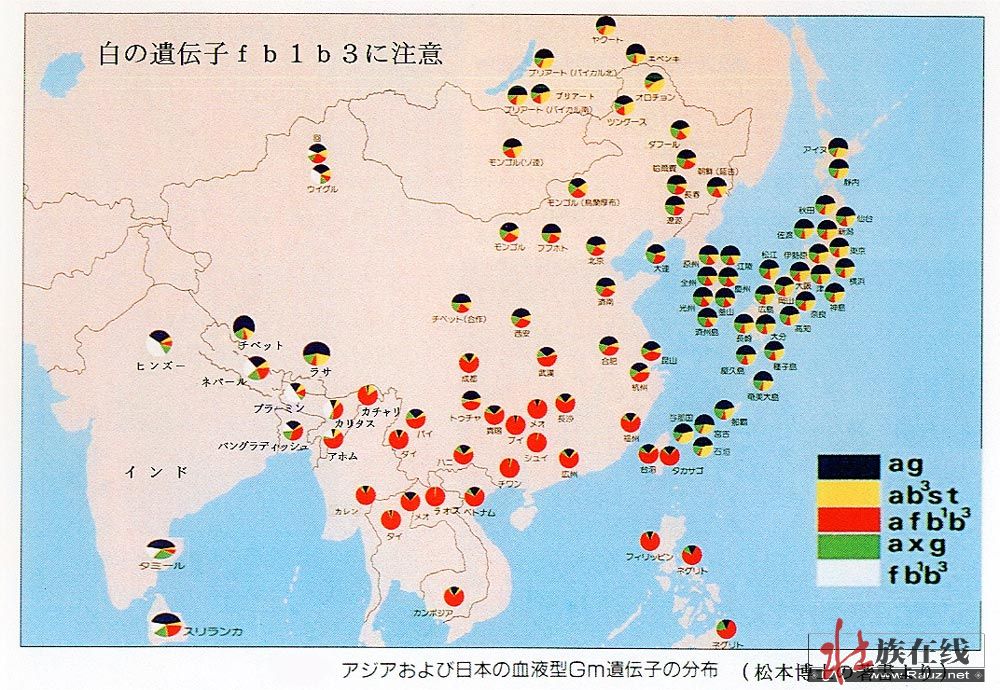
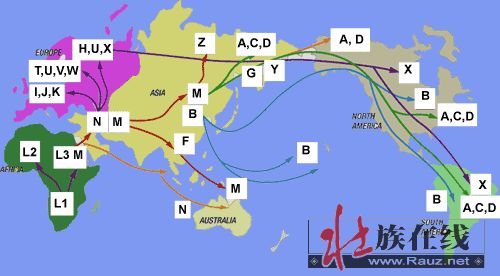
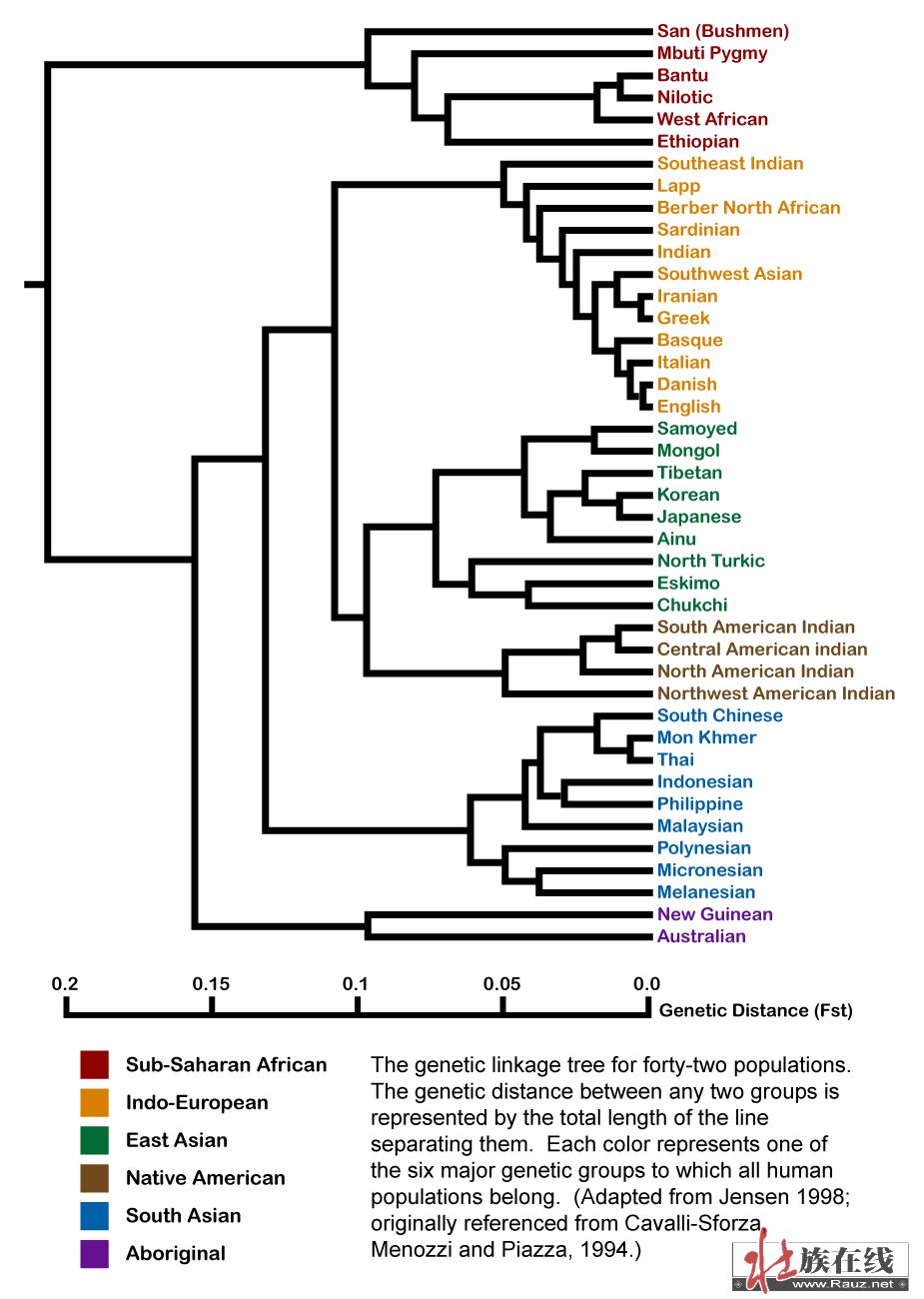
(转贴)2009-02-19 11:59:59)
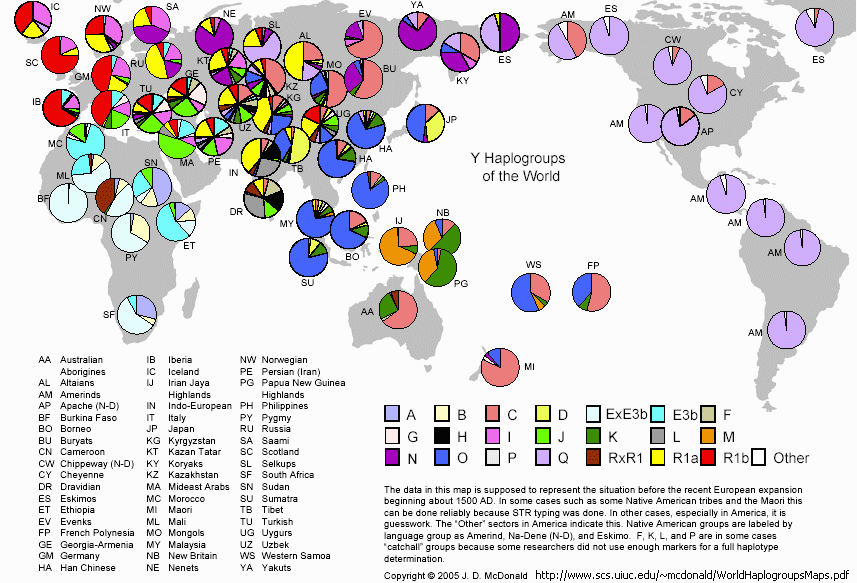
标签:y染色体单倍型 snp str 东亚 蒙古人种 黄种人 m175 汉族 汉藏语系 亚洲人 分类:基因谱系
一.古代东亚直立人的消失
东亚地区在10万——6万前,经历了一次剧烈的冰川期,这一时期,原来繁盛的东亚古代直立人(如北京人、蓝田人)消失了。在距离今天10——6万年前,东亚地区没有发现确切的人类活动遗址。
二.6万年前到达东亚的第一批古亚洲人——D(Yap)
距离11万年前,第一批智人走出非洲东部,到达今天的中东地区,他们的Y染色体标志为M168,今天,非洲以外的所有的土著民族都是M168人群的后代。
此后,他们不断向周边迁徙,大约距离今天60000±5000年前,他们中的一支终于到达东亚地区,当他们离开非洲时,他们Y染色体SNP为DE原始型,到达东亚时,他们的Y染色体SNP的标志已经发展成为D——Yap,而留在非洲的DE原始型则发展成为E,成为60%以上非洲黑人的类型。
但当时的非洲人绝不是黑皮肤的,而是黄褐色皮肤的,D人群也绝对不是黑皮肤,可能也是黄褐色皮肤的。
(在距离今天1100年前,漆黑色皮肤的西非班图黑人只分布在占非洲20%面积的西非地区,在非洲东部、中部和南部,占非洲面积60%以上土地上生活的是狩猎的黄褐色皮肤的布须曼人Bushman,今天他们仍然有很多人生活在南非,看上去肤色并不太黑,Bushman人兼具黄白黑人种特征,而且其Y染色体是最古老的A,可能是最古老的人类族群,直到大约在1000年前,黑色皮肤的黑人才扩散岛非洲东部、中部和南部)
当时的D人群的皮肤很可能就是和Bushman人一样的黄褐色,而且向北的过程可能还变的更浅了。
(所以称呼D人群为矮黑人并不准确,称呼老亚洲人更准确。)
D人群是最早到达了东亚的现代智人,但他们对现代东亚人的基因库gene pool贡献并不大。
D人群独占东亚地区的时间不超过1万年(6—5万年前),大约在距离今天5万年前,另一支老亚洲人——C人群来了,C人群是大部分东亚人的母系远祖。
D类型在东亚人中很少见。
D分为四个主要支系,D1、D2、D3、D*。
D1分布主要在华南地区,是年代古老的支系,历史超过5万年。D1今天的分布,向南进入东南亚半岛,如缅甸、泰国、马来西亚,向西进入青藏高原,在藏族中比例非常高,向北进入蒙古草原,在蒙古族的比例比较少(2/147),在汉族地区相当罕见。
D2只分布在日本,韩国有非常少的比例该成分,日韩之外完全没有D2。D2的历史大致有3万年左右,在日本土著阿伊努人Ainu中出现频率很高。关于阿伊努人有两组数据:一个是2006年Hammer的数据是D2为75%,C3为25%,另一个2002年Tajima的数据D2为87.5%,C3为12.5%。
D3分布在西藏,而蒙古地区也有部分D3,可能来自藏族的迁徙。
D*是D最古老的支系,有6万年的历史。低频率分布在西藏,在印度洋上安达曼岛土著人中出现频率100%,马来半岛的土著人群中也有低频率分布。但绝不能因此认为东南亚矮黑人等同于D或D等同矮黑人。
汉族中也有一定比例的D,主要是D1,少量为D*。如于敏2002年的数据,福建汉族D1为1/80,无D*,如Lu Chuncheng在2007年的数据,800个汉族D的出现10例,频率为1.3%(10/800),Hammer的2006年的数据,168个汉族中出现一例D1,为0.6%,Xue的数据中,166个汉族出现3例D,为1.8%,复旦在2000年的一组数据为362汉族出现1个D,为0.3%,另一组复旦数据里北方1054个汉族有22个D(2.2%),南方652个汉族中为9((1.4%),马明义2007年的四川汉族一文中,341个汉族有3个D*,8个D1,频率共计3.2%,其他一些数据里,汉族都没有发现D,出现频率为零,如Karafet在2001年的数据,Toru Katoh的2004年数据。总体看,D在汉族很罕见,但在甘肃和四川比例超过3%,可能是古羌族的影响(现代羌族D为18.2%)。
苗瑶人群中有一定古老的D1,显示了这些民族非常古老。D1在是瑶族中不同瑶族支系出现频率差异非常大,如复旦2002的数据里一支系瑶族(勉)为50%,但复旦2003年另一支系瑶族数据为0,Hammer在2006年数据里瑶族为1.8%(60人里仅1例,同论文中苗族为8.6%,58人里5个),而Xue的数据,Bama瑶族里有D而Liannan瑶族则没有D,可见,各支系瑶族D出现频率很不同,除了个别支系,总体上D频率并不高。
藏族中D是是最常见的类型,如Hammer的一组2006年的数据,105个藏族,D*为4人(3.8%),D1为16人(15.2%),D3为32人(30.4%),没有D2,D共52人(49.4%),又如Karafet在2001年的数据,75个藏族人中25个D*(含D3),12个D1,D共37人(49.3%)。今天,D和O在藏族大致各占半壁江山,藏族年轻的成分O和汉族的O在类型上一致,O3人群在大致5000年前从华北给青藏高原带来了汉藏语,而古老的D的显示了藏民族在人种上有悠久的历史,D人群进入西藏可能有2~3万年左右的历史。和藏族关系密切的普米族D比例高达72.3%(34/47)。
日本人是东亚另一个高频率D的人群,在如Hammer2006年的数据中,259个日本人发现90个D2,占34.9%,没有其他D类型。不同地区日本人中,D的分布很不平衡,在京都附近、九州岛、本州岛西部,是高比例的O而D少,而在北海道、种子岛和冲绳,D的比例则很高,显示了日本的最早的土著为D,后来为C,最后到达的是O人群,但O人群构成了当代日本大和族人的大部分主体父系成分,而日本土著阿伊努人则是D占80%以上,C占不到20%,没有O。
东亚其他人群D类型很少见,比如满族,在Karafet在2001年的数据里,复旦的2002年的数据,Toru Katoh的2004年数据里都没有发现D,Hammer在2006年的数据里满族也没有D,Han—Jun Jin的2003年的论文里44个满族里找到一个D,为2.3%,可以说满族人里D是相当罕见的。朝鲜人有一定比例的D,但比例不高,Toru Katoh的数据里延边朝鲜人D比例为2.5%(2/79),韩国人为3.5%(3/85),Han—Jun Jin的数据韩国人是2.5%(4/160),Hammer的数据韩国是4%(3/75),Wook Kim的数据是0%(0/216),Nonaka的数据是317个人里1个D1,12个D2,比例为4%。我国西北地区有一定的D,可能是古代羌族的遗留,如彝族人比较高,为16.3%(Hammer),现代四川羌族也比较高,为18.2%,西北回族也比较高,11.4%(Xue),9.3%(Karafet),9.2%(杨亚军),但在2001年spencer wells的中亚东干族里却没有D,Zerjal的2002年中亚东干人数据里也没有D。总体上说,现代西北回族的父系血统来源中,古羌族血统的影响是非常重要的来源。
D在东亚分布范围不大,在西伯利亚地区,当地北亚黄种人中并没有D类型,美洲印第安人中也没有D。
三.5万年前,C人群到达东亚,并构成现代一半以上东亚人的母系成分和黄种人体貌
D占据控制东亚的历史不超过1万年(距今6—5万年),在距今5万年前,C人群到达了东亚并控制了东亚。
早期的原始人渔猎生活,有三种主要的生活方式,一种是固定狩猎,一种是游动狩猎,另一种是就是海边食物采集。
固定狩猎的如尼安德特人(消亡了),游动狩猎的如克罗马农人(现在知道他们Y染色体属于I类型),而C人群,一般认为属于海边食物采集者。
海边食物采集者采集潮汐冲击到海岸上的海洋食物,每天都有固定的食物来源,他们可以一直沿着海岸快速移动,沿途都有稳定的高蛋白海洋食物,就这样,在5万年前他们迅速沿海岸线到达南亚、东南亚、东亚和澳大利亚。
C人群离开非洲时,还是原始的CF型,到达东亚时,已经变异成为新的C类型,而当时没有出非洲的CF原始型也在进化,后来发展成为F,后来又出现K,K在距离今天3万年前最终也进入了东亚。
C的最主要的几个支系为C*、C1、C2、C3、C4、C5。
C1只分布在日本地区的大和族人中(韩国也发现过一例),日本土著民族阿伊努人中主体是D2,少量C3,并无C1。日本人C1大致占5.4%,C3大致占3.1%(Hammer,2006年,259个样本),日本之外没有C1发现。
C2分布在印尼、巴布亚新几内亚等地,在印尼东部马来人中,C2占27.3%(Hammer,2006年,15/55人),新几内亚,86%(4/46),波利尼西亚人63.3%(37/60)。
C4、C5分布在南部印度人、斯里兰卡人和澳大利亚土著人中,超出黄种人的范围,不予以讨论。
C*是指除C1、C2、C3、C4、C5之外的C成分。
C*主要分布华南地区,如Hmamer数据中,瑶族为20%,土家为6.1%(另有C3为18.4%),苗族为1.7%(另C3为3.4%),马来亚为3.1%,印尼东部为5.5%(C2占27.3%),西部印尼为4.0%,维吾尔族为1.5%。C*可能主要分布在华南和东南亚地区,但实质上在华北地区也有很多。如XUE的数据里,如鄂温克为3.8%,赫哲族为6.7%,宁夏回族为11.4%,乌鲁木齐维族为3.3%,伊犁维族为2.6%,Bama瑶族为17.2%。可以说C*在东亚分布也是很广的。
对东亚人而言,真正重要的成分是C3。
C3大致产生于距离今天4—3.5万年前的东亚,并扩散到北亚和北美州、南美洲。
在如此广阔的范围内,C3的支系十分众多。
C3*、C3a、C3b、C3c、C3d分布区域各不相同,这种格局可能在3万年前就已经具有了雏形。
C3a只分布在日本和韩国,和D2、C1、O2b,还有母系的M7a一样,都是具有日本韩国特征的特有类型。
C3b分布在北美州西北部地区,是当地土著印第安人的特征,占当地印第安人的9%。
C3c是极其具有阿尔泰语系标志性的类型,在西伯利亚南部达到峰值,并深刻影响了周边的其他阿尔泰语系民族。
在西伯利亚的南部地区,C3c是最重要的成分。如Pakendorf在2006年的数据中,中部埃文人50%,东部埃文人61.3%,中部通古斯人为40%,北部说突厥语的通古斯人为15.1%,
Nyukzha通古斯人53.8%,Lengra通古斯人66.7%,Stony通古斯人70%,Okhotsk通古斯人为62.5%,Yenisey通古斯人为58.1%,可以说,在西伯利亚南部,尤其是通古斯人,C3c是最重要的成分。但在西伯利亚北部,N3则是最重要的成分,如雅库特人,N3占92.2%,而C3c才占1.1%,北部埃文人中,N3占90.9%,而C3c才占4.5,北极地区的爱斯基摩则以Q3和N3为主,没有C3c。在西伯利亚南部,N3的比例一般在20—40%之间,是南部西伯利亚通古斯人另一重要成分。
C3c在蒙古草原也有一定的分布,是蒙古语民族的标示性基因类型之一。如Toru Katoh的数据里,喀尔喀蒙古为15.3%(17/85),乌梁海蒙古为33.3%(20/60),Zakhchin蒙古为30%(18/60),Khoton蒙古为10%(4/40)。而Karafet在2001年的数据,149个外蒙古喀尔喀蒙古人,29个C3c,占18.1%。Xue的数据里内蒙古为8.9%(4/45),Zerjal在2002年的数据外蒙古人C3c为20%(13/65),spencer在2001年的数据里为45.8%(11/24)。Hammer的数据里,布里亚特人为4.9%(4/81),卫拉特人(阿尔泰)为5.1%(5/98),Pakendorf在2006年的数据,图瓦人(我国户籍登记为蒙古族),为9.1%(5/55),spencer在2001年的数据里图瓦人C3c为7.1%(3/42),Ivan Nasidze在2005年的数据,俄罗斯卡尔梅克人为37.4%(37/99)。综合看,核心的标准蒙古人一般有20%以上的C3c成分,其中内蒙古人最低,喀尔喀蒙古其次低,这两个群体低C3c主要是因为成吉思汗家族一簇的稀释和O3的高比例加入,内蒙蒙古人有最高比例的O3,超过了30%。外蒙古喀尔喀人也出人意外的有高比例O3,超过20%以上,而西部的图瓦人、Khoton人、卫拉特人、卡尔梅克人,O3都非常罕见,低于3%,甚至个别数据里根本没有O3,所以西部蒙古人可能原来本没有O3的,东部蒙古(喀尔喀—科尔沁—喀喇沁)则有高比例的O3,不同地区的蒙古人有不同的起源。
总体上看,突厥语各民族C3c并非主体成分。只有哈萨克族C3c特别高,如spencer在2001年的数据里57.4%(31/54),Zerjal在2002年的数据63.2%(24/38)。同样spencer的数据里,吉尔吉斯人为7.7%(4/52)卡拉卡尔帕克人为2.3%(1/44),7个地区共计326个乌兹别克人里一共4个C3c(只分布在其中2个人群,另5个人群无C3c)。中亚维吾尔人、土库曼人、喀山鞑靼人、克里米亚鞑靼人、阿塞拜疆人里没有C3c。Zerjal的数据里,中亚维吾尔人3.3%(1/33),吉尔吉斯人为12.2%(5/41),其他突厥语民族乌兹别克人、土库曼人、阿塞拜疆人没有C3c,Cengiz的2003年土耳其人数据,523个土耳其人只有5个C3和1个O3,其他都非东方草原血统。总体看,突厥语人中,哈萨克人与其他所有突厥人都明显不同,他们可以说与草原和西伯利亚南部边缘的蒙古人基因上非常接近,所不同的仅仅是他们讲突厥语而非蒙古语,吉尔吉斯人也有非常高比例的东方草原民族血统,尤其接近萨彦岭——阿尔泰山等地的土著游牧狩猎民族。其他突厥民族的主体成分来自中亚农业土著居民,与哈萨克、吉尔吉斯等草原游牧民族差异极其明显,而N3占92.2%的雅库特人与其他突厥语民族基本没有血统相似性,是被同化的地道的北极边缘的土著人。
C3c在我国东北其他阿尔泰语居民中也有一定分布,Hammer数据里,鄂温克为34.1%,鄂伦春为68.1%,辽宁满族没有C3c,Xue的数据里,鄂温克为26.9%、鄂伦春族为41.9%、赫哲族为11.1%、锡伯族为4.9%,岫岩满族为2.8%,达斡尔族为2.6%,Toru Katoh的数据辽宁满族为7.9%。总体上看,东北地区各阿尔泰语系民族父系远祖主体来自华北平原地区的东亚人群,受到西伯利亚居民的影响的主要是鄂伦春和鄂温克人,其他人都和南部西伯利亚边缘的通古斯人关系甚远。
C3c在阿尔泰语周边的汉、韩、日民族并不存在。如2001年spencer的北方汉族、韩国人中,2004年Toru Katoh的辽宁汉族、延边朝鲜人、韩国人、日本人,2006年Hammer数据的华北汉族、华南汉族、台湾汉族、韩国人、日本人、冲绳人,2006年Xue数据里的哈尔滨汉族、伊犁汉族、兰州汉族、成都汉族、梅县汉族、延边朝鲜人、韩国人、日本人中,都没有发现任何一个C3c,所以可能情况是,C3c主要是随着阿尔泰语系的扩散而扩散的,周边的中、日、韩人里没有受到这种扩散的影响。
C3d是C3各支系中的另一个非常重要的支系,C3d主要分布在汉族、越南等民族中。
C3d在汉族中数量不高,但非常古老。实际上,整个C3家族,也是越向南越古老,STR多样性越高,显示了C3整体上是从华南地区向华北再向北亚扩散的。
C3d(C3*)在今天很多汉藏语民族中都有高比例分布,如Hammer数据中的土家族,为18.4%,如复旦数据的白族,为16.7%,汉藏语人的C3和北亚人的C3并不相同,相反,汉族、越南两族人的C3在STR上十分接近,同时和白族、土家族的C3d十分接近,有相同的起源,可见汉族和南方越民族的关系十分接近,独特的C3d显示了汉族等众多汉藏语民族共同的南方起源性质。
C人群对应的母系为M系(M*、M7、M8、M9、M10、C、D、G、Z)。今天,东亚人母系成分中,M对应了大致60%的成分,北方略多,南方略少。从常染色体角度上可以说,今天大部分东亚人都有一般以上古亚洲人的血统。
部分人尤其是一些网友推测,C人群,尤其是C3人群,外貌上已经完全是现代黄种人外貌。称呼“棕色人种”并不一定准确。是否确实如此,在没有论文数据情况下,我们不好定论。但我们要承认,C人群对现代东亚人基因库gene pool的贡献并不逊色于N和O人群,唯独只是在Y染色体上略少一些而已。
四.东亚人主体的父系远祖——O人群
东亚真正的主体是NO人群。
NO是K的一个分支,大致在3.5万年前进入,并在3万年前左右分化成N和O两个姊妹类型。
NO到达东亚时,东亚地区已经为C3所占据,但NO类型在很短时期内就成功的取代了C3,成为了东亚的主人。
NO是最原始NO原始人群的类型,出现在以下人群中:达斡尔族2.6%、鄂温克族3.82%、赫哲族2.2%、回族2.9%,华北汉族2.3%,华南汉族2.5%,越南人3.1%,外蒙古人0.7%、布依族5.7%、日本人2.1%、韩国人2.3%、彝族人2.3%,Bama瑶族2.9%。
N、O的分布在黄种人差异巨大。对东亚黄种人而言,O比N多得多,而对北亚黄种人而言,只有N而无O,对于泰国以南的黄种人而言,只有O而无N。
O类型分为很多类型,主要有O*、O1、O2*、O2a、O2b、O3等。
O*在是O的原始类型,在东亚只是零星的低频率找到,比如Hammer和XUE的数据中,共有以下民族中找到过最古老的O*:满族5.8%、壮族5.0%、外蒙古人0.7%、藏族1.0%、达斡尔族2.6%、鄂伦春族3.2%、兰州汉族2.9%、黎族5.7%,在lu chuncheng在2007年的论文中,在800个江苏汉族中,找到11个O*,比例为1.4%。在马明义2007年的数据中,341四川汉族中找到4个古老的O*,出现频率为1.2%。
NO和O*两个古老的类型发现频率都很低,而其他的O类型也比较常见,下面一一介绍。
O1根据李辉2007年研究,是良渚文化的主体类型,同时,根据金力李辉的数据,河北磁山文化的主体也是O1,同时红山文化中也存在部分O1。
O1今天汉族中的分布如下:在lu chuncheng在2007年的论文中,在800个江苏汉族中139个O1,占17.4%,体现了良渚文化对现代江浙人仍然有很大影响,虽然主体已经变为龙山系统的O3。在马明义2007年的数据中四川汉族10.9%(341人中的37人),复旦2002年的数据为北方汉族1054人有36人,频率3.4%,南方汉族652人有100人,频率为15.3%,Hammer的数据,168个汉族(44个北方,124个南方人)有16人,频率9.5%。Toru Katoh的辽宁汉族为4.8%,Zhou Ruixia的2007年数据,甘肃汉族为3.4%(3/87),李辉2007年闽越一文中,195个福建汉族(南澳30、莆田36、汕头68、潮州22、闽南37、闽东2)共找到13个O1,比例6.7%,同一论文中河南安阳为4%(2人/50人)。
总体上看,汉族北方地区大致只有5%以下的O1,而南方地区一般有10%以上的O1,显示了古代良渚文化队华北汉族的血统影响很小,而对南方汉族却有一定的影响,尤其是对长江下游地区。
2003年福建长汀的148个客家人中只发现4个O1,比例只有2.7%,却找到110个O3,比例达到74.3%,客家人O1明显比周边南方汉族低,却和北方汉族高度一致,不但显示了客家汉族父系血统起源的北方特征,也从一个方面证明了古代时期的北方汉族和现代的北方汉族是一致的,的确没有太多的O1成分。
汉族以外的东亚地区,O1的比例如下:(Spencer,2001)东干人5.0%、吉尔吉斯人5.8%、外蒙古4.2%、韩国人4.4%,(karafet,2001)韩国2.7%、满族3.8%、鄂温克族2.4%、宁夏回族1.9%,(Toru Katoh,2004)满族3.0%、韩国人2.4%、日本人2.6%,(石宏,2006)白族6%、基诺族4.4%、傈僳族6.1%、壮族4.2%、布依族4.3%、土家族7.4%、普米族4.2%,(Hammer,2006)台湾原住民89.6%、土家族8.2%、苗族6.9%、瑶族1.7%、越南5.7%、壮族10%、马来西亚6.13%、菲律宾10.4%、东部印尼12.9%、西部印尼20.0%,(Xue,2006)回族8.5%、达斡尔族5.1%、鄂温克族7.7%、锡伯族7.3%、满族2.9%、黎族26.5%、羌族15.2%、畲族14.7%,Bama瑶族2.9%,Liannan瑶族5.7%,(Zhou Ruixia,2007)藏族7.9%。
总体上看,O1最高频率出现在马来语居民中,台湾土著最高,其次为中国华南地区的各民族,如黎族、土家族、羌族、畲族等。东北的一些少数民族也有比较多的数量,显示O1曾经分布在从华南到华北再到东北地区的广阔地区,但华北地区的O1则在后来O3崛起后消失了,华南和东北地区的则保留了。
O2*是扣除O2a和O2b的原始型O2,在东亚也有比较多的分布。
O2*分布在以下人群中(karafet,2001):华北汉族4.5%、宁夏回族3.7%、韩国人1.7%、满族人5.8%、鄂温克族4.9%、鄂伦春族4.5%、台湾汉族7.1%、土家族4.1%、彝族2.3%、瑶族20.0%、外蒙古人1.3%,(Xue,2006)回族2.9%,赫哲族2.2%,达斡尔族2.6%、满族8.6%、内蒙古人2.2%、鄂伦春族6.5%,哈尔滨汉族8.6%、伊犁汉族9.4%、延边朝鲜4.0%、羌族2.9%、黎族2.8%、bama瑶族37.1%、外蒙古人1.5%。
综合看,东亚人中,华南的瑶族人两组数据,一个37.1%一个20.0%,比例都很高,其他民族都很低,但东北地区也有比较高的比例,显示O2*最初可能是连在一起的,但中部被一个其他人群割断了,推测可能是龙山文化(O3)的崛起取代了O2*,造成了O2的隔断。
O2a,是O2的最大的一个支系,高频率的出现在东南亚地区,尤其壮侗语(Daic)居民中。
2007年李辉的史前长江一文显示,江西吴城文化中,O2a是主体成分,今天,O2a也是主要分布在长江以南的汉族之中。
O2a在汉族中,主要出现在南方,如文波2006年的数据中,甘肃汉族1/60、河南汉族0/50、辽宁汉族1/48、内蒙汉族1/60、山东汉族2/185、陕西汉族1/90、安徽汉族0/22、湖北汉族0/18、江苏汉族4/100,总体上长江之北的北方汉族为9/633,只有1.5%,复旦2002年的数据,北方汉族1053人中13人为O2a,比例为1.35。总体看,O2a在北方汉族大致也就是1%的比例而已。而长江之南的汉族中O2a则非常高的频率,如2006年数据中,浙江5/106、上海汉族0/55、江西1/21、福建长汀客家4/148、广东汉族12/64、广西汉族7/26、湖南汉族2/15、四川8/63、云南汉族3/93,591人中发现42例子,比例7.1%,而复旦2002年的数据中,南方汉族,652人中发现47人,为7.2%,两者比例十分接近,可以说,大致长江之南O2a也就7%,只有广东18.8%,广西26.9%偏高,,扣除两广之后的南方汉族,文的文章中O2a大致为4.6%(29/501),这个比例显然与北方汉族是十分接近的,显示了除了两广外,汉族内部成分的高度一致性。
汉族之外,O2a高度集中在南部壮侗语居民和周边民族中,如复旦2002年数据中,壮族10/28,泰国13/20、高棉族9/26,又如董永利2004年的数据,傣族44/112、壮族18/47、水族8/40、布朗族14/28、傈僳族20/49、纳西族19/40、布依族28/48,XUE的数据里,布依族19/35、哈尼族17/34、黎族21/34,2006年石宏彝族中的僰人为16/32。Han—jun Jin的数据,越南为5/50、泰国为26/55、印尼人为10/36、菲律宾为1/77。
O2a在北方民族中较少出现,只有达斡尔族是例外,比例高达6/39,鄂温克族也有O2a,XUE数据为3.8%(1/26)、Hammer数据为2.4%(1/41)。石宏的僰人一文,289个藏族中有4个O2a,和北部汉族的比例很接近。大致长江以北,O2a很少(除达斡尔、鄂温克族)。
有观点说日本人中存在大量从东南亚北上的南岛语人口,甚至网络上有说法日本天皇家族的成员检测为O2a,我认为这个不太可能,日本人中O2a很少见。Xue的数据里日本为2/47、Hammer的数据日本为5/259,Han—jun Jin的数据为1/108、Nonaka的数据8/729,总体上,为16/1143,比例非常小的,不太可能有大量南岛人参与了日本人的形成。
O2b是东亚人又一常见类型,在东亚地区主要分布在日本和韩国,但在东南亚,则有更广阔的分布。
(XUE的数据,2006):日本13/47、韩国12/43、延边朝鲜族7/25、锡伯族1/41、满族2/35、赫哲族2/41、达斡尔族1/39,(Hammer的数据,2006):日本29.5%(76/259)、满族3.8%(2/52)、韩国36%(27/75)、越南4.3%(3/70)、印尼16%(4/25),Han—jun Jin的数据:日本25.9%(28/108)、朝鲜19.4%(31/160)、印尼19.5%(7/36)、菲律宾1.3%(1/70)、越南14%(7/50)、泰国5.4%(3/55),总体看,在日韩和东南亚地区,O2b分布很广,在汉藏语居民中,O2b基本是没有的,在阿尔泰语系中,主要分布在东北地区,蒙古和突厥语地区基本没有。
O3是东亚的主人,也是汉族的主体。
根据2007年研究,山西陶寺遗址的龙山文化居民中,上层贵族100%为O3类型,而根据2008年金力李辉的数据,陕西仰韶文化的居民的Y染色体SNP类型也是O3,所以,我们可以清晰的看出,古代中原居民,尤其是贵族,都是O3占有绝对主体的,仰韶文化和龙山文化居民的Y染色体成分是基本相同的,种族上是同一成分。
O3是汉族的绝对主体,占据非常重要的位置,根据2004年文波的数据,山东汉族为59.5%(110/185)、河南52%(26/50)、陕西汉族56.7%(51/90)、云南汉族63.4%(59/93)、广东汉族53.9%(34/64)、四川汉族57.1%(36/63)、浙江汉族是50.9%(54/106),张咏莉2002年福建汉族的为58.8%(47/80),2006年Hammer的数据北方汉族65.9%(29/44)、台湾汉族49/84(58.3%)(注意这个数据和张咏莉2002年福建汉族的58.8数据几乎一样,类似的还有Nonaka的台湾汉族,183个里106个O3,比例58.2%,福建闽南汉族和台湾汉族数据高度一致),XUE的2006年数据,哈尔滨汉族O3频率是68.6%(23/35),Lu chuncheng的数据中南京汉族为53.6%(429/800)、马明义的数据中四川汉族61.8%(211/341)2007年Zhou ruixia的数据,甘肃71.3%(62/87),所有数据中,除了河北人83.3%(10/12)由于样本太少外,目前看到的O3出现最高比例的汉族人群是客家人,根据2003年李辉的数据显示,福建长汀客家人中O3的比例高达74.3%(110/148),还有一个类似的数据,是2002年李辉在《广西六甲人来源》一文中的数据,为77.8%,(21人/27人)。
总体上看,所有汉族中,O3都是主体的,不同的取样可能造成数据有波动,但总体出现频率大部分一般在50-70%之间,虽然幅员辽阔人口众多,但汉族内部却表现出了惊人的一致,显示了汉族的共同起源。
今天10亿汉族的绝大多数,都是5000年前中原原始居民的直系后代。主体是仰韶文化和龙山文化的O3(两种文化居民的父系成分相同),O3也就是来自中原仰韶文化和龙山文化的居民构成了各地汉族父系远祖的压倒性绝对多数,其次是河北的磁山文化的O1,红山文化的O3、O2和O1,和良渚文化的O1,其次还有湖北大溪文化和江西吴城文化的O2a,今天广阔的中国土地,仍然为5000年前中国远古土著居民的子孙所占据。
汉族的平话人群是汉族唯一的例外,其O2a超过O3而占主体,与其他汉族不同。
东亚其他民族中,O3也非常常见,这并不等于说这些民族中有汉族血统,因为O3至少在1万年前就扩散到了整个东亚,他们中的O3的一部分就是这些远古人的后代,而非全部是后来中原统一崛起的汉族带去的。
其他汉藏语居民:spencer2001年数据,东干族40%(16/40),zerjal数据2002年:土家族52.8%(26/49),董永利的数据55.8%(38/68),和汉族非常接近,实际很多汉藏语居民都和汉族接近,又如白族2004年董永利数据为50%(25/50),而2004年文波的数据为50.8%(31/61),白族和土家族可能和汉族有非常紧密的民族起源关系。而其他汉藏语民族和汉族关系要相对远些,如XUE的数据:羌族30.3%(10/33),藏族40%(14/35)、布依族31.4%(11/35),如董永利2004的数据:哈尼族17/41、景颇族4/17、彝族12/41、纳西族1/41、布依族2/48(和XUE的数据显著不同)、普米族4/47。
中国境内其他民族的数据还有:傣族21/112、黎族1/31、壮族8/28。
在东亚的其他地区,如日本韩国,2006年hammer数据日本为20.2%(52/259),韩国为40%(30/75),Toru Katoh的数据,延边朝鲜29.1%(23/79)、韩国34.1%(29/85)、日本16.2%(19/117),XUE的数据:延边朝鲜40%(10/25)、韩国17/43(39.5%),Nonaka的数据韩国为44.5%(117/317),wook kim的数据韩国47.2%(102/216),总体上看,韩国O3比例大致在40%以上,为第一大类型,日本大致在20%,在日本O3比O2b少。
在阿尔泰语系中,东北地区满语族人O3比例非常高,为满族、锡伯族、赫哲族的第一大类型。在蒙古族人群中,内蒙和喀尔喀外蒙人最高,大致在20-30%之间,西部蒙古O3非常少,基本都在5%以下或者根本没有。西伯利亚地区没有O3的分布。突厥语人中,O3不高,但分布很广,最高为裕固族和撒拉族,大致30-40%,维族大致10%,越向西则越来越少,哈萨克族大致在10%,乌兹别克大致2%、而到了土耳其地区,523个土耳其人只有一个O3,只占0.2%。
O3占各阿尔泰语人群的具体数据如下:
满语族:UE数据:满族37.1%(13/35),Hammer数据满族38.5%(20/52)、Toru Katoh数据满族42.6%(43/101),文波的满族数据为44.4%(8/18),XUE中,赫哲族为44.4%(20/45)、锡伯族26.8%(11/41),鄂温克族23.1%(6/26),鄂伦春族19.4%(6/31),hammer鄂温克族数据为24.4%(10/41)。
蒙古人:喀尔喀蒙古zerjal数据22.8%(34/149),卫拉特蒙古1.0%(1/98),spencer数据外蒙古8.3%(2/24)、Toru Katoh数据喀尔喀蒙古18.8%(16/85)、乌梁海蒙古6.7%(4/60)、zakhchin蒙古8.3%(5/60)、khoton蒙古2.5%(1/40),Xue的数据内蒙古28.9%(13/45),外蒙古10.8%(7/65)、达斡尔族25.6%(10/39)。
突厥语人:Zhou Ruixia数据:裕固族34.2%(18/52),维族12.2%(6/49),zerjal2001年的数据维族10.5%(7/67),XUE的数据:伊犁维族15.4%(6/39),乌鲁木齐维族6.5%(2/31),spencer在2001年的数据特别丰富:吉尔吉斯1/52,哈萨克9.3%(5/54),中亚维族12.2%(5/41)、图瓦人1/42,卡拉卡尔帕克人11.4%(5/44),克里米亚鞑靼人1/22,乌兹别克人14/326。Pakendorf在2006年的数据里,图瓦人1/55。土耳其人1/523。可以说,突厥人中,O3是普遍存在的,但比例都不高,和突厥邻近的塔吉克、伊朗、巴基斯坦等地也发现了O3,可能是突厥人、蒙古、契丹人或者汉唐时期华人的影响。尤其是哈萨克维吾尔等人群中O3比例远远高于西部蒙古人,显示了O3的来源绝对不只蒙古草原,古代中原人也参与了这些民族的形成。
五.走出东亚的土著人群——N和Q1
N和O是姊妹类型,大约在3万年和O分化,并在距今2~1.5万前从东亚出发,进入西伯利亚和东欧。
但N遗留在东亚的部分仍然是很多,不容忽视。
N可以分为N*、N1、N2、N3四个部分进行陈述。
N*是汉族的常见类型,在10个汉族人群总数1475人的样本中,共有100个N,占6.8%,最低频率为0,最高频率为15.0%(Hammer华南汉族6/40),其余如南京6.1%(49人/800人)、华北9.1%(4/44)、台湾6.0%(5/84)、四川8.8%(30人/341人)、兰州6.7%(2/30),所以说N是汉族非常重要的一个来源,以往很多数据没有单独检测该SNP,所以全部被归类到了K里,造成了汉族有K*的假想,实际上,汉族是不含东南亚的K*和中东的K2的。
N*最高频率出现在彝族等西南民族中,彝族Hammer2006年的数据为30.2%(13/43),附近的土家族4.1%(2/49),而四川汉族中为8.8%(30人/341人),西南地区可能是N*的大本营。
N1分布要小的多,目前看,最主要分布在中国,如Hammer的数据里为华北汉族2.3%(1/44),韩国人1.3%(1/75)、满族人1.9%(1/52),Xue的数据里为满族5.7%(2/35),锡伯族2.4%(1/41),布依族5.7%(2/35)。由于检测该SNP的论文只有这两篇,所以目前发现的人群还很少。
N2分布在西伯利亚西部和乌拉尔山脉地区,在蒙古地区有低频率分布。
如Pakendorf的数据中,图瓦人27.3%(15/55),由于该SNP也和N1一样很少被检测,所以很多数据都被归类到了K中。
在西伯利亚的通古斯人(Evenki)中,N2也很常见,如stony通古斯人为27.5%,yenisey通古斯为9.3%,中部地区的通古斯人为16%。
在蒙古和东北地区,如zerjal的数据,外蒙古为6.0%(9/149),中国鄂温克族2.4%(1/41)、布里亚特2.5%(2/81),如XUE的数据,内蒙为0/45,外蒙为2/65,东北地区,赫哲族最高,为8/45、满族为2/35、鄂伦春族为2/31,可以说N2在我国北方少数民族中也有一定分布,但汉族没有找到过任何N2。
N3是N的最重要的支系,分布地区从中国一直延伸到北欧的广阔地域。
在欧洲人中,在芬兰人、爱沙尼亚等人群达到60%,在立陶宛、拉托维亚等人群中占50%,在俄罗斯北部人中占35%,中部占20%,南部占10%,占挪威和瑞典人的5-10%,占德国人的3%。这里我们不过多讨论N3在欧洲的分布。
N3最高频率分布在西伯利亚,kharkov和brigitte的数据里,293个雅库特人有270人是N3,占92.2%。在通古斯人(Evenki)中okhotsk通古斯为37.5%、yenisey通古斯为9.7%、lengra通古斯为22.2%、中央通古斯为18%、讲雅库特语的通古斯人为70.7%、nyukzha通古斯人为20.5%、北部埃文人为90.9%,中部埃文人为8.3%、南部埃文人为12.9%,可以说N3在西伯利亚北部占绝对主体,南部西伯利亚则N3减少,C3c占了主体。
在蒙古地区,Pakendorf的数据,图瓦人的N3为9.1%(5/55),spencer的数据图瓦人为21.4%(9/42),外蒙古人,zerjal的数据N3为2.0%(3/149),XUE数据为内蒙13.3#(6/45)、外蒙古7.7%(5/65),toru Katoh数据喀尔喀蒙古4.7%(4/85)、乌梁海蒙古8.3%(5/60)、zakhchin蒙古3.3%(2/60)、khoton蒙古0(0/40),derenko布里亚特蒙古为18.9%(45/238),zerjal布里亚特为28.4%(23/81),卡尔梅克蒙古ivan的数据为1.0%(1/99),Xue的数据达斡尔族的N3为7.7%(3/39)。
在东北地区通古斯语人群,Toru Katoh满族为0(0/101),XUE的数据锡伯族为(2/41),赫哲族0/45、鄂伦春族0/31、鄂温克族0/26、满族为0(0/35)、hammer的数据,满族为0(0/52),鄂温克族为0、鄂伦春族为1/22。基本上,N3对东北地区通古斯语民族的影响非常微弱。
在汉族、朝鲜人、日本等东亚民族中,基本没有N3。但应该注意到的是在spencer的东干人中有N3(1/40),在zerjal的西北回族也有N3(1/54),显示回族有部分来自北亚草原的血统。
Q1是指汉族特有的M120,在2008写做Q1a1。
曾经有观点认为Q1是汉族特有的类型之一,但这种观点正在受到挑战。
应该说,目前公布的绝大部分数据里的Q1都来自汉族(目前论文数据里找到的大致在300个左右,有270左右来自汉族),复旦2002年的数据,北方汉族1054个人有60个Q1,占5.7%,南方汉族652人中有10个Q1,占1.5%。如马明义的四川汉族数据,341个人中有9个Q1,占2.6%,lu chuncheng的南京汉族中,800个汉族有24个Q1,占3.0%,Zerjal的数据中,北方汉族Q1为4.5%(2/44),台湾汉族1.2%(1/84),Nonaka的台湾为1.1%(2/183)。
在汉族以外的汉藏语人群中,Q1也有发现,如董永利2005年白族起源一文中,在白族6.7%(2/30)、土家族3.1%(1/32)、藏族2.2%(1/46),如文波在摩梭人一文中,普米族为2.1%(1/47)。可以说,在汉族之外,Q1也是存在的,主要分布在西南汉藏语居民中,Q1不应该简单的看成汉族独有。
汉族以外发现Q1,东亚之外在东干人、巴基斯坦hazara人中找到过该类型。在东亚,在中国之外,在日本kanto地区为0.7%(1/137),冲绳2.2%(1/44),nonaka数据韩国0.6%(2/317),zerjal数据越南7.1%(5/70)。
目前比较有说服力的观点认为,Q1是1.5~1.0万年前进入东亚的细石器文明的创造者,他们可能是从中亚进入草原后进入东亚的,并且一直分布到整个东亚,直到大致1万年前,O各个支系的扩张,才冲淡和驱逐了他们,造成了他们只有残留在汉族和其他东亚人群中,频率非常低。
六,综论
对东亚人来说,O是最大成分,O在除了西伯利亚的所有人群都有分布,而且在东亚和东南亚所有人口超过1000万的大人群中都是最大的成分(且均超过50%)。
C是其次大的,尤其是对北方人群来说,在蒙古和西伯利亚通古斯人中都超过了50%,但除了蒙古和哈萨克外,所有C占主体的民族人口基本都不超过10万,很多都是游牧和狩猎的几千人组成的小部族。
N是第三位的,但对中国西南的一些民族和西伯利亚,也是非常重要的。
D的分布比较局限,只有日本和西藏有比较高的频率。
Q1只低频率出现在东亚,没有一个人群超过10%。







 发表于 2009-6-29 00:57:00
发表于 2009-6-29 00:57:00

 QQ空间
QQ空间 腾讯微博
腾讯微博 腾讯朋友
腾讯朋友 微信
微信 转播
转播 淘帖
淘帖 分享
分享 收藏
收藏 支持
支持 反对
反对 发表于 2009-6-29 19:10:00
发表于 2009-6-29 19:10:00

 发表于 2009-6-30 00:19:00
发表于 2009-6-30 00:19:00