Paternal Genetic Affinity between Western Austronesians and Daic Populations
Li Hui, Wen Bo, Chen Shu-Juo, Su Bing, Pramoonjago Patcharin, Liu Yangfan, Pan Shangling, Qin Zhendong, Liu Wenhong, Cheng Xu, Yang Ningning, Li Xin, Tran Dinhbinh, Lu Daru, Hsu Mutsu, Deka Ranjan, Marzuki Sangkot, Tan Chia-Chen, Jin Li (2008)
英文原作下载:http://li-hui.net/Documents/A41.pdf ,以下是我的节选翻译。
南岛西部人群和壮侗人群父系的紧密遗传关系
摘要
背景:南岛语是在东南亚、大洋州和印度洋地区扩展最广的语系。以他们语言的相似性为基础,这个语系包括了马来-波利尼西亚和台湾土著语。语言上的相似性还导致了有争议的台湾是南岛语原乡的假说,在民族学家、语言学家、考古学家和遗传学家中都有争论。普遍公认东部南岛语人(密克罗尼西亚人和波利尼西亚人)源自西部南岛语人(东南亚岛屿和台湾原住民),而且大陆上的壮侗族群则被假定为所有南岛语人的源头。
结果:在报告中,我们研究了30个中国壮侗族群,23个印度尼西亚和越南的马来-波利尼西亚族群,共1509个Y染色体的非重组区的20个SNP位点和7个STR位点。这三个群体在父系遗传谱系方面显示出许多共同之处,混合分析证实了壮侗族群在遗传方面几乎未受中国汉族影响,而且构成了印尼人的遗传的主体。O1a*在大部分族群的样本中高发,而在其他语系族群中十分少见。,O1a*单倍群的网络图解释了印尼人的世系并非如语言学研究结果的那样源自台湾原住民,而是来自壮侗族群。
结论:我们发现,与台湾原乡假说相反,东南亚岛屿族群在父系遗传谱系上并非台湾起源。另外,我们的还发现台湾原住民和印尼人在父系上很可能都源自壮侗族群。这两支人群似乎都经历了独立发展的过程,我们的研究结果显示遗传上支持一个包括台湾原住民、壮侗族群和马来--波利尼西亚族群在内的大族群。
结果
为确定壮侗人群和南岛西部人群的遗传关系,我,我们测定了23个东南亚岛屿族群和11个台湾原住族群(群体所在地见图1,族群信息见表1),1509个Y染色体的非重组区的20个单核苷酸多态性位点和7个短串重复序列位点。本次研究囊括了几乎全部中国壮侗族群和台湾原住族群的样本。
另外,现已完成了对包括所有东亚和东南亚语言集团在内的134个东亚族群SNP位点定义单倍群的主成分(PC)分析。结果显示语其他东亚和东南亚群体相比,壮侗人群最为接近西部南岛族群(图2),表明壮侗人群和西部南岛族群有很近的遗传关系。壮侗-东南亚-台湾集团从其他种群中的分异与主成分2有高度相关性(详见附件1)。此外,O1a-M119是台湾原住民的优势单倍群(平均77%),检测值从54%至100%不等(见表2,O1a*和O1a2之和),但其他东亚族群中则不然(<5%)。因此,预计O1a-M119能为勾画壮侗族群和西部南岛人群的关系提供丰富的信息。
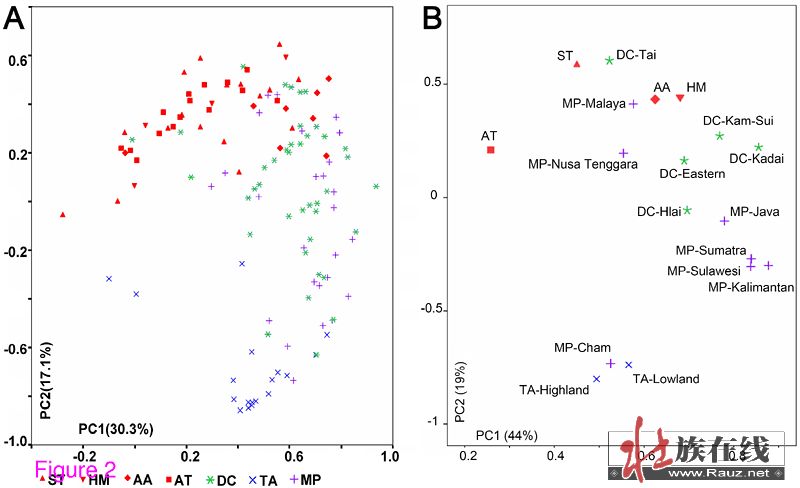
图2中的主成分图显示一些壮侗族群接近汉藏族群的簇。可能是因为壮侗族群和汉藏族群有共同的祖先,可能导致他们在遗传上有相似之处。然而,对此也可以有另一种解释,就是东亚大陆上的壮侗族群在遗传上可能受到过中国汉族的影响,因为他们从2500年前就开始比邻而居。混合分析可以评估假定现代壮侗人群的壮侗或汉族祖先的比例,而一些被隔离的壮侗族群可以作为混合分析中的父系族群。海南的原住族群(黎族、加茂人和仡隆人)和台湾原住岛民可以假定为相对孤立的,因为他们的文化很少受到大陆上的外来文化的影响。所以,在遗传结构上这些原住岛民可能很类似于壮侗祖先。
为评估假定的大陆壮侗族群中的汉族遗传影响,我们将大陆壮侗族群、海南原住民、台湾原住民和中国汉族的Y-SNP数据应用于混合分析中。为了分析,我们将后3个群体合起来视为大陆壮侗族群的父系群体。结果显示海南原住民的遗传贡献非高(2.145±0.927),然而中国汉族(-0.314±0.422) 和台湾原住民(-0.831±0.662)的贡献几乎没有。这里的ADMIX程序算出的遗传贡献负值暗示对现代壮侗族群没有贡献的可能。这个结果显示壮侗族群的父系遗传谱系是相当清晰的,壮侗人群与西部南岛人群之间的遗传关系几乎未受族群混杂的影响。
东南亚岛屿族群可能也是混合的。在我们的研究中,假设东南亚岛民在父系遗传上是是三个群体的混合:壮侗人群、台湾原住民,以及很接近巴布亚人的巽他群岛土著居民。我们完成了印度尼西亚人的混合分析,并在分析中加入了从文献中得到的巴布亚人数据作为父系族群结构之一。我们的分析显示如下的混合比例:壮侗群体(0.713±0.124),台湾(0.143±0.125)和巴布亚人 (0.144±0.050),表明壮侗祖先对印尼人的父系遗传贡献是最强势的。在这些数据中存在一些不确定性,因为我们关于东南亚岛民是混血的假设是无法得到检验的。
正如O1a*是壮侗和西部南岛人群最具特色的单倍群,我们使用O1a*个体的7个STR位点评估了壮侗人群、印尼人和台湾原住民之间的遗传距离,(遗传距离见图3,STR原始数据见附件2)。我们的研究显示台湾和印尼族群之间遗传距离是最大的,几乎差不多是壮侗群体和台湾原住民遗传距离的3倍。壮侗群体和印尼人之间的遗传距离很接近壮侗人群和台湾原住民的距离。这些发现表明印尼人和台湾原住民在遗传上都与壮侗族群的更接近,超过了这两个西部南岛群体之间的距离。此外,在O1a*个体的7个STR位点上的多态性在壮侗语人中比在印尼人和原住民中更高发(图3)。多态性最高的族群未必总是最古老的,也可能是与周边族群混血的结果。但是,壮侗语人O1a*单倍群较高的多态性应该是族群年代最古老造成的,因为这种单倍群在周边人群中几乎不存在,所以不是混血导致了更高的多态性。我们所取得多态性和分歧时间结果显示,壮侗人群很可能是印尼人和台湾原住民父系遗传世系上的祖先群体。其他的Y单倍群(如O3-M122、O2a-M95)也呈现与O1a*相似的图谱,显示壮侗人群与印尼人和台湾人的遗传距离比后两个族群之间的距离更接近(表3)。有趣的是O2a甚至可能进一步追溯到南亚语人群,正如近期研究所提出的观点。
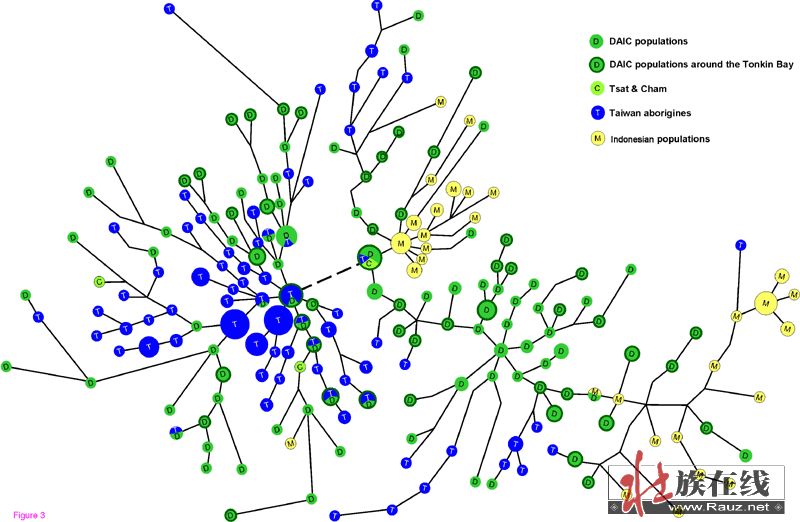
我们用三个人群O1a*个体的7个STR突变位点构建了中接网络(图3)。假如东南亚岛民的台湾原乡假说(THH)是真的,也就是说东南亚岛屿个体来源于台湾原住民,那么网络中的东南亚世系和台湾世系就应该有共享节点和(或者)连线。在图3中,壮侗世系(绿色节点)构成了网络的中心。所有的东南亚世系(黄色节点)和台湾原住民世系(蓝色节点)直接或是间接地共享或连接着壮侗世系。相反地,没有台湾世系(除了一个)分享或者连接到东南亚世系的节点。这些发现表明东南亚岛民并非直接源自台湾原住民,而应该是东南亚岛民和台湾原住民各自独立地起源于壮侗群体。
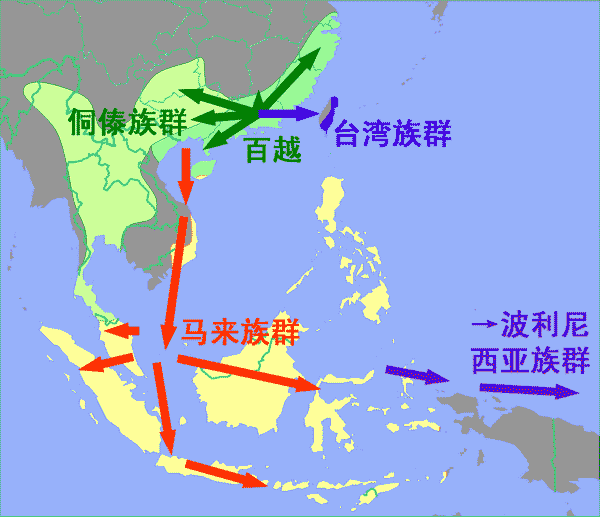
我们进一步注意网络中那些与东南亚世系相连的壮侗族群世系。有趣的是,大部分与东南亚相连的壮侗单倍型不是来自海南,就是来自海南西北面的广西(图3中带深绿外圈的绿色节点)。这些海南和广西族群均位于北部湾周边区域(即东京湾,Gulf of Tonkin)。特别是越南南部的属于马来-波利尼西亚族群的占人(cham),以及海南的回辉(Tsat),可以看到他们在网络中联系着壮侗人群和印尼人。所以我们假设东南亚岛屿族群起源于北部湾周边地区,他们穿过印支半岛到马来半岛向南迁徙,而后再扩散到太平洋和印度洋的大部分岛屿。
我们估算了网络中O1a*单倍群的年代。总体年代是33765±5221年,与末次冰期相吻合。网络中所有壮侗系样本的年代是33193±5577年,与O1a*的总体年代很接近。由于台湾与壮侗的单倍型大范围交叠,很难估计台湾簇的真实年代。有些交叠表明发生过从壮侗族群到台湾原住族群的多次移民。我们估计网络中左侧那个台湾簇的有14659±3110年。所有台湾样本的总体年代估值为21268±3148年。有趣的是,后面这个年代值很接近台湾已发现的最早的人类遗存,即左镇人。由此我们推断O1a*个体从大陆向台湾岛的移民出现在旧石器时代。
由于在网络中可以观察到两个清晰明确的东南亚岛屿单倍型的簇,我们计算了两者的年代。左边的东南亚簇的年代是9895±2393年,可是右边的簇却达25880±7137年。语言学上对马来—波利尼西亚语的起源年代的估计比我们的估值要年轻得多,大约5000-6000年前。此外还可发现在网络中到壮侗单倍型和东南亚岛屿单倍型有一些交叠,说明在东南亚人脱离壮侗祖先群体向外移民过程中可能发生过瓶颈效应。地理学上的瓶颈效应可能来自狭窄越南海岸。所以O1a*单倍群很可能在马来—波利尼西亚语7500年产生之前由此向东南亚岛屿扩散。但是晚近时期O1a个体向东南亚岛屿移民的可能性是不能忽视的,因为遗传学时间估计的精度不足以排除这种可能性。
应当注意的是,在“特别快车”假说中存在两种不同的角度:(1)移民的起源,也就是台湾原乡说,以及(2)移民模式,也就是从印尼开始的快速扩散。在研究中,为验证台湾原乡说,我们检测了先前研究中很少涉及的包括壮侗语人和东南亚岛民在内的西部南岛人群。我们发现台湾不象是东南亚印尼人的原乡,至少不是其父系主体世系的故乡。尽管台湾原住民和印尼人都源自壮侗人群,但两者父系主体世系并非单一起源表明其分化是相互独立的事件。
有意思的是,我们的研究表明家猪在东南亚群岛和太平洋地区的分布与西部南岛族群分布基本一致。台湾及远到密克罗尼西亚的家猪都直接来源于东亚大陆,而那些东南亚群岛和波利尼西亚的家猪是来自印支半岛。假设家猪是被人类群体在早期迁徙中引入的,那就意味着人类同样可以顺着这两条的路线抵达东南亚和太平洋岛屿。
事实上我们的检测结果与澳泰语群单一起源理论相一致,澳泰语群由壮侗语、马来—波利尼西亚语和台湾土著语组成。我们的检测结果表明,在这次研究加入壮侗人群和东南亚岛屿人群对研究南岛人群的起源是绝对必要的。如果不加入这些群体,波利尼西亚和台湾原住民会显得相互十分接近,从而导出所有南岛人都是起源于台湾的结论。
我们的结果表明北部湾极有可能是东南亚岛屿族群的原乡。鉴于人类群体从印尼东部向太平洋岛屿移民过程的复杂性,以及西部和东部南岛人群在遗传方面的显著区别,我们在研究中没有选择波利尼西亚的数据。作为替代,我们只分析了西部南岛群体。波利尼西亚族群缺少O1a-M119的事实是令人好奇的,而且不能简单地以瓶颈效应来给出波利尼西亚人Y染色体单倍型表现出高多态性的原因。
与我们对父系遗传谱系的发现相一致,对马来半岛群体的线粒体DNA研究也表明,大约在末次冰期(Last Glacial Maximum)时原始马来人的祖先居住在印支半岛。后来这支祖先群体经由马来半岛扩散到东南亚岛屿。对东南亚岛屿人群的mtDNA研究也表明,即使发生过一次南岛语人从台湾的移民,在人口统计学上也是一个次要的事件。
我们的大部分结论都是基于对O1a*的分析,一个在这些族群Y染色体谱系中只占少部分的单倍群。O1a*谱系的频率在台湾族群中显著高发,不过在马来—波利尼西亚人和壮侗人群中并不特别多。一些族群事件可能还涉及到其他的Y染色体谱系。有其他次要的、不同来源的父系谱系也很正常,比如说南岛群体到来之前的印尼土著族群,还有更晚近来自南亚的移民。东亚和东南亚人之间遗传关系的复杂程度超出了我们的预料。
结论
我们的结果显示,在父系遗传谱系上壮侗人群比其他东亚民族群体更接近西部南岛族群。O1a-M119是壮侗族群和西部南岛族群中主要的Y染色体单倍群,其STR多态性表明西部南岛的两个群体——台湾原住民和东南亚岛屿土著,各自独立地起源于壮侗群体。因此,东南亚岛屿族群很可能起源于北部湾地区——壮侗民族的原乡,然后经越南走廊迁徙至印度尼西亚。与此相反,台湾原住民族是从中国大陆直接移民。我们的研究结果表明从遗传上可以推断出一个包括台湾原住人群、壮侗族群和马来—波利尼西亚人在内的大群体。
粗略看了一下,不求甚解,也看不太懂,呵呵。
表呢?
这篇论文的确艰深了点!不对照李辉之前的一些论文不容易看懂。我等会把他最近另一篇关于海南黎族起源的文章也贴出来,以便对照理解。
文中的表格和附件都很大,请直接下载原文查看。
Paternal Genetic Structure of Hainan Aborigines Isolated at the Entrance to East Asia
Li Dongna, Li Hui*, Ou Caiying, Lu Yan, Sun Yuantian, Yang Bo, Qin Zhendong, Zhou Zhenjian, Li Shilin, Jin Li (2008)
英文原文下载:http://li-hui.net/Documents/A40.pdf
隔离于东亚入口的海南原住民父系遗传结构
要点:为评估孤立的海南岛遗传结构和南方入口处最早的遗传结构,我们研究了全部6个族群的405个海南原住民Y染色体,他们受到新近大陆人群变动和混杂的影响很小,这里我们报告在海南原住民中O1a*和O2a*单倍群是占优势的。此外,大陆上占优势的单倍群O3在原住民中的频率很低,说明他们是孤立的群体。聚类分析性表明海南原住民在2万年前被隔离,在两种主要的单倍群进入东亚之后(3.1-3.6万年前)。
结果和讨论
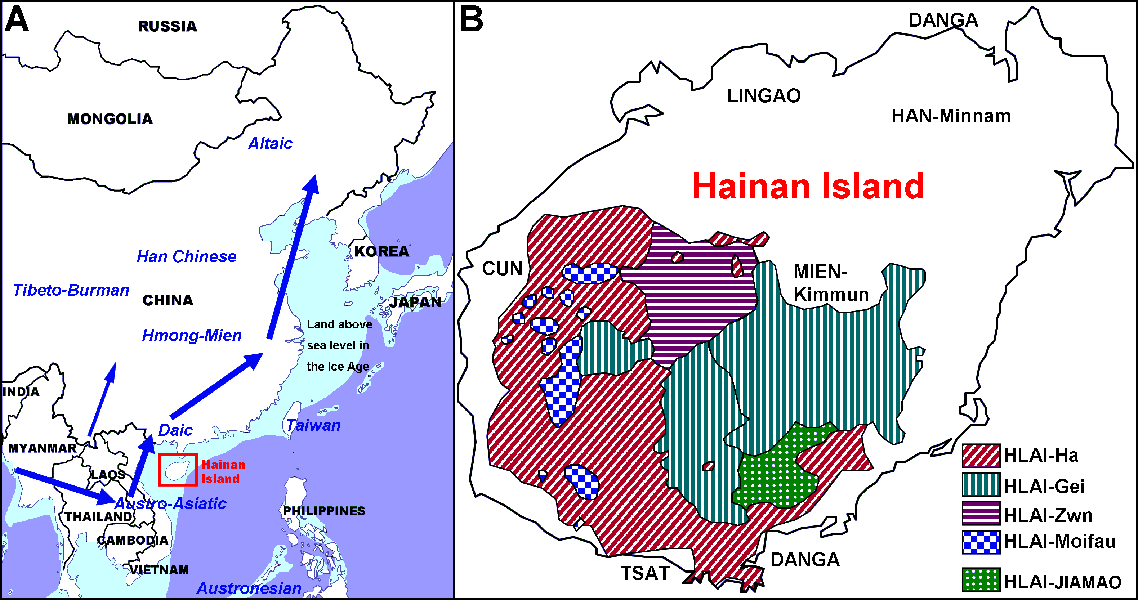
在我们的研究中,我们分析了海南原住民6个人群的Y染色体差异。我们检测了405个海南男性个体的22个单核苷多态性(SNPs),核7个短串重复(STR)多态性,并确定了基于YCC和ISOGG标准命名法的单倍群。我们建立了在Y-SNP单倍群频率方面非常近似的族群取样。O1和O2是在各人群中出现最多的,而且很可能是海南原住民的原始单倍群,在海南原住民之一的杞黎中,这些单倍群的总频率达到了100%。表明这是一个显著瓶颈效应的人群。事实上,杞黎分布在偏僻的山区,而且是由一个很小的人群发展起来的,杞黎Y-STR多态性较低支持了这个观点。O1和O2都是台湾原住民和东亚最南部常见的单倍群。在中国东部和南部沿海、婆罗州和菲律宾的南岛语人群中O1和O2也高发,他们的语言由语言学上近似的壮侗语和南岛语组成。与壮侗族群相比,其他族群的O1比例相对较低,特别是在印支半岛西部的操南亚语的土著人群中。O3是汉藏语族群中共同的单倍群,汉族(50.51%),藏缅族群(54.70%),在海南原住民中是很少的(6.91%),虽然在台湾原住民中的频率达11.36%(0%-37.6%),以及在大陆南方(壮侗19.6%)和中部(孟高棉54.02%)原住族群中有较高的频率。这表明与台湾和大陆南方土著族群相比海南群体与汉族的男性遗传混杂较少。还有与大陆族群不同,D、P、N、Q等单倍群未出现在海南原住民中。
壮侗语和南亚语族群是散居于大陆南方并交错分布。我们假定在他们之间必然有充分的基因流动,而且可以被归入一个群体,这与他们的Y-SNP频率的情形是相似的。黎族亚群构成了南部群体的外部分支,而且其中的瓶颈和被隔离的黎族杞黎,更是处于这个分支的外侧。这个结果显示黎族完全不同于大陆上混杂的族群。在北方群体中,孟高棉、中国汉族和藏缅系人群相互之间是最接近的,虽然在遗传上南岛语和阿尔泰人群的全部影响都来自汉藏语系群体。我们找到了这个群体中的外部分支是台湾原住民和海南仡隆人,呈现出相对较低的单倍群差异。但是还不清楚为什么发现仡隆人和台湾原住民接近。
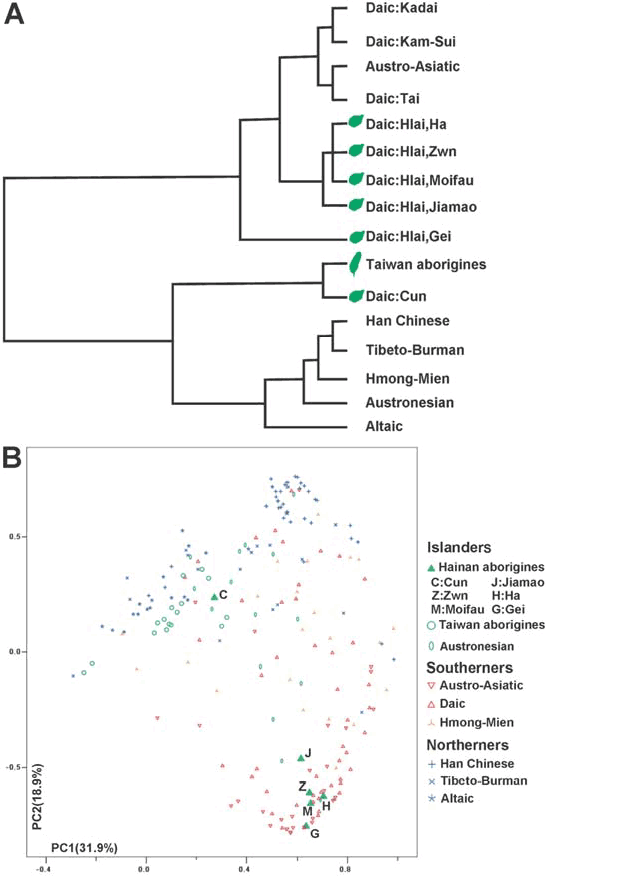
主成分分析也显示南方和北方的族群是分开的。在主成分图的基础上,台湾原住民接近北方族群,而海南原住民是分为两支不同的群体。仡隆人可以与台湾原住民一起被放到北方族群中。所有的海南黎族群体非常接近南北分布的南部边缘,再次验证了他们孤立的遗传结构。这种分化尤其准确地显示了杞黎是处于南部边缘的位置。
依据SNP分析,海南原住民与北方族群是隔离的,特别是在华北人群向南方移民时携带着高频的O3单倍群。孟高棉族群向南的移民可能也为O3在华南的频率的增加作出了贡献。我们最近的研究显示孟高棉和汉藏语族群最近其的共同祖先在中国西南部地区。正如我们提到的,海南原住民中缺少O3。但是海南原住民占优势的单倍群O1和O2,同样也在大陆壮侗族群和台湾原住民中占优势。单倍群SNP分析不排斥海南原住民和壮侗-台湾人群之间的基因流动。
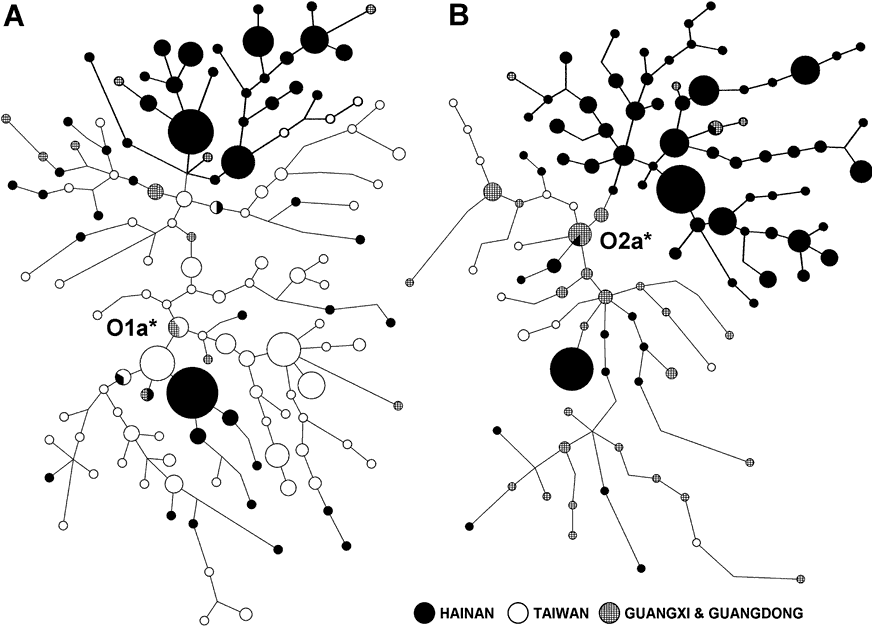
在这个网络中,涉及的数据是来自台湾原住民,以及海南北面的两个省份广东、广西的壮侗语人群。在图3网络的每个顶端,海南原住民(显示为黑色节点)从其他族群中很少的个体中形成了几乎独立的分支(粗线显示),暗示海南原住民是从其他壮侗人群与台湾原住民中隔离出来的。此外,黎族大部分的单倍群位于顶部的分支,虽然仡隆人的单倍群在两者的网络中形成了两个很小的分支(在网络较低的部分的黑色节点)。黎族分支的尺度相对较大,几乎占据了网络的三分之一。在O2a*的网络中,黎族的分支比大陆和台湾的部分更大,说明黎族是比大陆壮侗族群和台湾原住民更古老,而不是源自大陆壮侗族系的群体。从STR来看这些大尺度的分支是通过相当长的时间所变异和形成的。
我们估计人群年代为:O1a*约3.6万年,在O1a*网络中海南分支的年代大约1.9万年,而O2a*的年代是3.2万年,在O2a*网络中海南分支的年代大约2.6万年(表2)。请注意我们对O2a*的估值是基于华南人群,因此不同于世界范围内O2a*的年代。这两个单倍群同样很少出现在华北,在华南人群中这两个单倍群的年代很可能就是他们进入东亚的时间。O1a*和O2a*的年代很接近,下限分别是1.8万年和2.6万年。这个时间框架与末次冰期高峰期相吻合,这时中国海洋的大陆架在海平面以上而且对现代人进入东亚来说提供了一个捷径。
对于早期迁徙进入东亚的现代人来说可能存在一条以上的通路。另一种可能的进入东亚的途径是从东南亚-东亚的西部接壤(从缅甸到云南和中国内地,图1A)。我们假定O3单倍群是经这条西部路线由汉藏和孟苗祖先群体带入的,尽管还没有足够的证据(我们最近对苗瑶和孟高棉人群的研究给出了可能存在西线的线索)。然而,O3也可能是通过相同的一条东部路线出现的,但是晚于O1a*和O2a*出现的时间。O3单倍群后来通过重新分布影响了很多其他人群。根据O3在海南岛孤立族群中很少来判断,当壮侗人的祖先首次进入东亚时,没有O3从海南岛附近进入。无论如何,可以确定最早沿东线进入东亚的移民携带的O2a*是一个最古老的单倍群(超过4万年,金力未公开的数据)。O2a*单倍群可能也是最早抵达海南岛的人群。海南岛上的O2a*分支的年代已确定约为2.6万年前,这个数据比海南最古老的三亚落笔洞遗址更久远。我们提出这就是海南原住民被隔离的年代。将来可能会在海南岛发现更早的考古遗址。
我们的O1a*网络分析图显示台湾节点比海南节点更接近网络的中心,表明在地理上台湾很接近这个单倍群的起源地。因此,O1a*可能起源于海南岛以东区域,而后倒流回海南,可以确定海南O1a*分支比O2a*分支更年轻。O1a*同样可以在5000年前的中国东部沿海的新石器时代的样本中找到。东亚的新石器时代始于8000年前。O1a*也有随着新石器文化扩散到海南岛的可能。O1a*在海南的年代(约1.8万年前)比海南的新石器时代遗址(约6000年前)更古老。一种可能的解释是这个年代实际计算的是在到达海南岛之前,O1a*人群离开O1a*祖先群体(可能是台湾原住祖先)的时间。我们计算出台湾和海南原住民O1a*之间的分歧时间是2.2(95%置信区间1.25-4.68)万年。
总之,我们的发现揭示了海南原住民来源于末次冰期进入东亚的早期移民,并从那时起就与世隔绝。海南原住民几乎没有受到东亚大陆上人群重新分布的影响,海南原住民的Y-SNP图谱最接近最初的遗传结构。我们认为海南原住居民不仅能揭示东亚人的起源以及他们的遗传特色,而且也可以当作东亚人群遗传研究的模型。
我们知道,隔离的人群对一些遗传学研究来说是很有用的。例如,复杂病症的致病因素显然在隔离人群中可以被简化,更便于分析。此外,相对较大的族群和久远的历史使得大多数海南原住民不太可能发生遗传漂变,却可以发展出与低Y-SNP多态性相对而言更高的Y-STR多态性。可以确信海南原住民也有着比其他东亚人群更高的常染色体变异多态性,更低的连锁不平衡(linkage disequilibrium),这在疾病关联性研究中可以消除由高连锁不平衡引起的错误的阳性结果。因此,我们认为在许多遗传学研究中都可以将海南原住民作为东亚隔离人群的模型。
艰深呵.看得头都晕菜了.
O1起源于广东,O2是南亚系?
深奥!看不太明白。
但估计让某些人不高兴!
O1起源于广东,O2是南亚系?
没错!现在的研究结果就是如此。O1起源于广东,而后O1a产生于更东部的区域,靠近台湾。
记得刚开始的时候01a是被定义为百越染色体,而O2a被定义为苗瑶,O2为百濮,现在看来,其实O1a和O2a染色体都跟壮侗族群有着密切的联系
我想可能应该这样理解:O1a其实是原始南岛人群的标志性单倍群,它产生时百越还未诞生。百越族群实际上是原始南岛人群的大陆支系同化其他族群所形成的民族-文化共同体。文中已经清楚地说明壮侗族群中的O2a最初是来源于南亚语人群。
海南岛原住民的O3染色体会不会是在迁至北部湾之前的东南亚就已经携带有?
文中的意思是,进入海南岛的早期成分是O2a和O1,没有O3。早期O3主要沿“缅甸—云南”从西南地区进入东亚,因此原始的O1a在回迁到北部湾的过程中混杂O3的可能性不大。
Pinghua population as an exception of Han Chinese’s
coherent genetic structure
Gan Rui-Jing, Pan Shang-Ling, Mustavich Laura F, Qin Zhen-Dong, Cai Xiao-Yun, Qian Ji, Liu Cheng-Wu, Peng Jun-Hua, Li Shi-Lin, Xu Jie-Shun, Jin Li, Li Hui*: the Genographic Consortium (2008)
英文原文下载:http://li-hui.net/Documents/A38.pdf
平话人群是汉族一致性遗传结构的例外
(节选翻译)
摘要
汉族是世界上最大的单一民族群体,由十种汉语方言支系构成。平话支系是个例外,这个群体的遗传结构已被广泛研究,Y染色体和mtDNA数据证明了所有汉族人群遗传结构的一致性。因此,尽管分布在壮侗系和苗瑶系民族比汉族人数更多的广西自治区,但我们相信平话人群这个汉族中的早期分支应该也不会例外。我们研究了470个平话群体的个体样本(其中包括195名男性)以及来自广西壮族自治区北部六个地区(贺州、富川、罗城、金秀、三江和武宣)其他的族群(壮族、侗族、仫佬、拉珈和瑶族)。对这些样本的mtDNA和Y染色体进行了检测。在平话人群中发现了高频的Y染色体单倍群O2a和O*,这两种单倍群总是高频出现在南方少数民族中。只有罗城县和金秀县的平话群体保持着汉族常见的O3a5a。在平话人群中找到的mtDNA谱系有B4a、B5a、M*、F1a、M7b1和N*,呈现出类似周边原住族群尤其是壮侗民族的遗传图谱。平话人群、其他汉族支系以及其他东亚族群的遗传分支分析(系统树、主成分分析和STR网络),表明平话人群比其他汉族支系更接近南方少数民族。混杂分析同样证实了这个结果。在语言、文化和自我认同方面,平话人群的祖先被汉族所吸收,从而导致平话人群成为汉族一致性遗传结构的例外分支。
材料与方法
族群样本
我们采集了来自广西壮族自治区的贺州、富川、罗城、金秀、三江和武宣县的197个平话人群的样本。血液样本随机选自健康和无关的个体,包括101名男性和96名女性。这些样本分别取自居住在不同乡镇的个体以避免取样偏差。临近的对照样本同样选自广西原住少数民族,包括拉珈人、壮族、仫佬族、侗族和瑶族。对照样本也来自健康和无关的个体,样本数273个(94名男性和179名女性)。(以下略)
结果
Y染色体单倍群频率
195个男性个体的Y染色体单倍群是根据ISOGG分类法确定的。数据来自我们新近采集的样本,在表1中给出。平话人群主要的单倍群是O2a*、O3a5a和O*,接近于周边的少数民族的遗传图谱(O2a*、O3*、K和O*)。但是汉族的高频单倍群O3a5a似乎只在罗城和金秀两组较小的样本中有较高的发生频率。和之前研究的结果一样,O3a5a在原住人群中是比较罕见的。一些单倍群的分布与种族有关:O1a和O2a*在壮侗人群中高发;而O3*和O2a*在苗瑶人群中高发;O3、O3a5和O3a5a在汉族中较常见。O2a*在平话人群中发生频率最高,说明该群体更接近南方少数民族而不是其他的汉族群体。
mtDNA单倍群频率
在470个样本我的ID发现了43种mtDNA单倍群(表2)。平话人群中高发的mtDNA单倍群是B4a、B5a、
M*、F1a、M7b1和N*,这很类似周边少数民族的发生频率,尤其是壮侗族群。金秀的平话群体有高频的F3单倍群,有别于其他群体。罗城的平话人群常见M7*单倍群,频率也比其他人群高。这两组人群的mtDNA和Y染色体单倍群图谱都有别于其他平话人群,但是也可能是样本数较少的缘故。B、F、R9a、R9b、N9a和M7单倍群是南方原住族群常见的单倍群。壮侗族群的特征单倍群是B4a、B5a、F1a、M7b1、M7b*、M*、R9A和R9b,苗瑶的的特征单倍群是B4a、B5a、M*、M7b*、C、B4b1、M7b1、F1a、B4*和R9b,汉族则是A、C、D、G、M8a、Y和Z。平话人群的mtDNA单倍群图谱与南方原住民族的图谱更接近,而与汉族有一定差异。
系统树聚类分析
在图2的系统树中,我们对多个东亚族群的Y染色体和mtDNA数据进行聚类分析。在分析时合并了属于相同语群的群体。十个汉族分支和六个其他语群以族群平均发生频率进行合并。
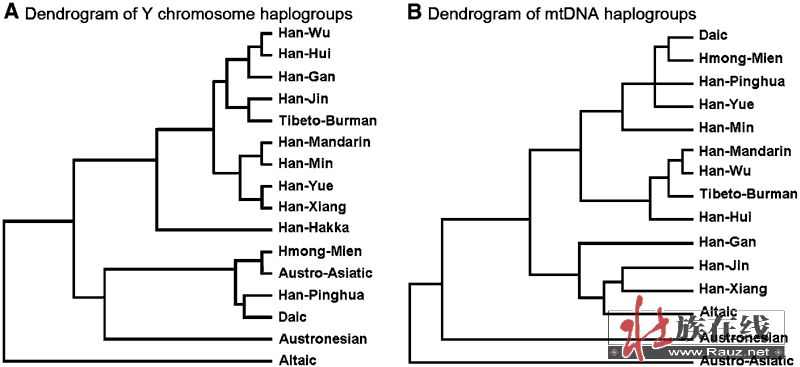
在Y染色体单倍群的系统树中,除平话人群以外,所有汉族分支聚为一个组,表明汉族群体父系遗传结构具有高度一致性。藏缅族群落入了汉族的簇中,这与汉藏语群的分类地位相吻合。南方原住民族语群构成了另一个簇。而平话人群的分支落入了这个聚类中,显示了平话人群的南方特征。北方主要的语群——阿尔泰语系,独立于上述两个聚类之外。在汉藏聚类簇的内部,人群主要是按地理位置而不是按语言聚类。在地理上接近的湘语和粤语分支同样在聚类树上也最接近,尽管在语言上湘语应该属于吴、徽、和赣方言的语支。在mtDNA数据的聚类上地理的影响表现得更为清晰。最南方的群体(壮侗、苗瑶、平话、粤、闽)全部聚为一个簇。
主成分分析
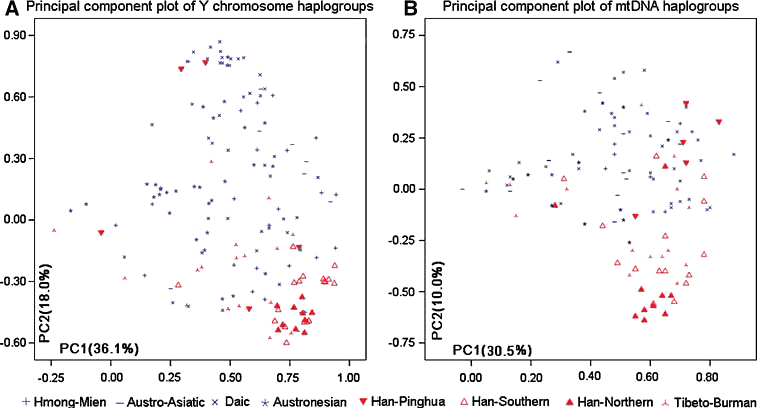
在平话人群的主成分分析中用到了汉族和少数民族的数据。在图3a的Y染色体散点图中,可以看到在主成分1和主成分2上南方和北方汉族群体都紧密地聚集成一个簇,在图3b的mtDNA散点图中,北方和南方汉族仍然紧密聚集,只有少数南方汉族群体出现离散。因此,主成分分析反映出汉族高度一致的遗传结构,而南方少数民族数据的分布普遍很分散。在Y染色体散点图上,除了金秀和罗城的群体外,其他平话人群都是很远离汉族的聚类簇。在mtDNA的散点图上,所有的平话群体都离散于南方少数民族之中,也反映出族群之间基因流动上的性别差异。母系遗传谱系总是在父系支配婚姻下非常频繁地流动。因而平话人群的母系遗传谱系才会比父系更接近他们周边的民族——壮侗和苗瑶族群。
主要单倍群的STR网络分析
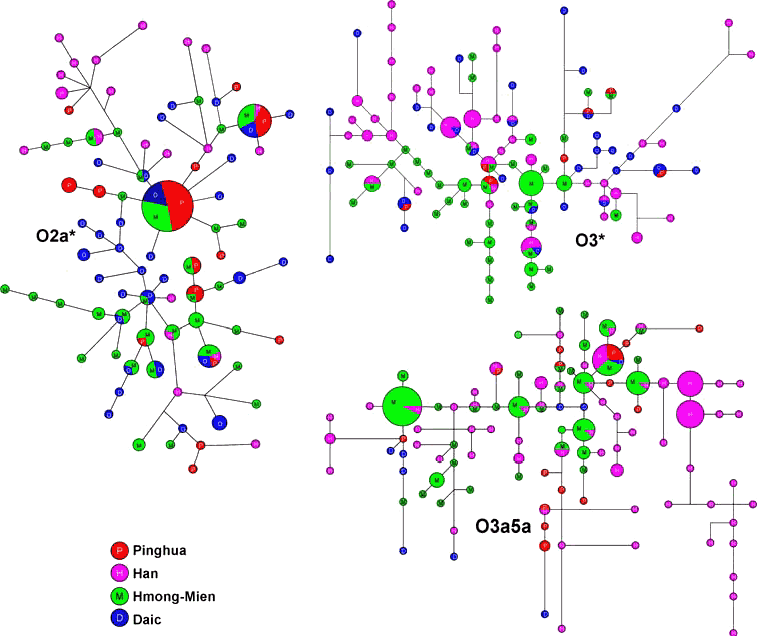
Y染色体单倍群的主体是O2a*、O*、O3a5a。在参考族群中,我们只发现在壮族中有高频的O*;而在汉族群体大量可用的数据中,O*很罕见或者未测出。因此我们无法分析O*的网络结构。在图4中,我们分析了O2a*、O3*和O3a5a单倍群的6个短串重复序列多态性(STRP)特征,并将平话群体样本的STRP单倍型与汉族、壮侗族群和苗瑶族群进行比较,发现O2a*网络中至少存在两个核心单倍型:一个在图上部,而另一个在近中间的位置。上方的分化中心为壮侗、孟棉和平话群体占据。绝大多数其他的汉族分支在图上半部分,可以看到一个汉族特有的小分支存在于粤东闽南语人的末端节点中。这个汉族小分支源自苗瑶族群的单倍型,并与相邻的汉族和畲族(东部苗瑶族群)的历史相吻合。纯粹的壮侗族群或者苗瑶族群同样可以观察到,这表明STR网络提供了与种族有关的信息。大多数壮侗和苗瑶民族的单倍型位于O2a*网络的中心。而大多数平话及其他汉族群体很可能源自这个单倍型,因为他们大都位于网络的末端。平话人群与南方少数民族共享的单倍型更多一些,而与其他汉族群体共享的较少。他们也有很多单倍型与南方少数民族相连,表明大部分携带单O2a*单倍群的平话人来源于南方少数民族。如果与两大原住族群比较的话,平话人的单倍群与苗瑶族群有着更密切的联系。在O3*的网络中,苗瑶单倍型更接近中心,汉族的小分支出自苗瑶的单倍型,而壮侗的单倍型处于外围。平话个体和壮侗个体共享更多的单倍型。在O3a5a的网络中,平话个体与汉族个体的关系最为密切,这些样本都来自罗城,在平话群体中罗城的样本具有最高频率的O3a5a。
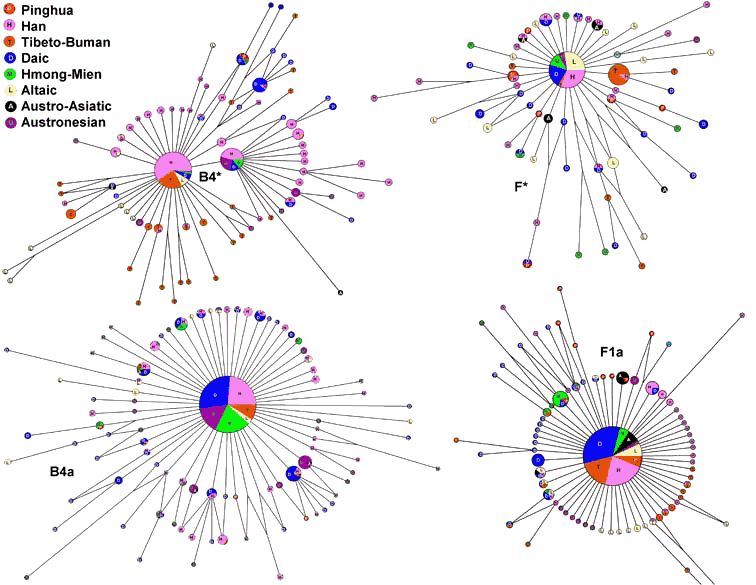
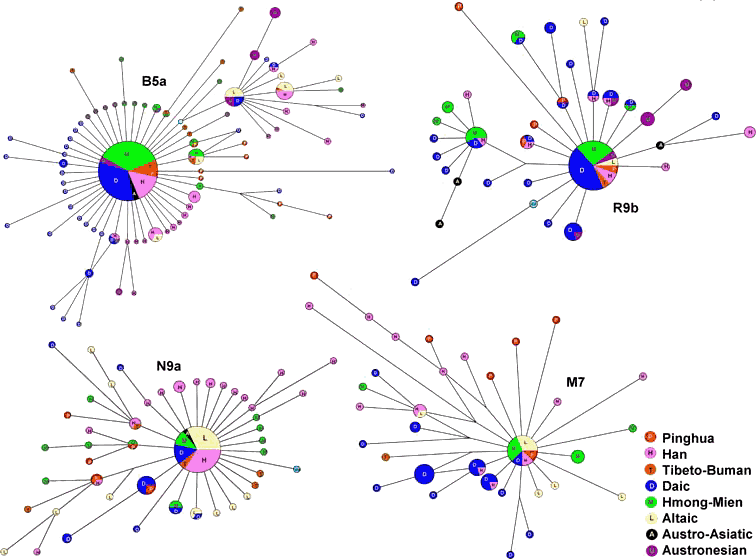
mtDNA的HVS-1(高可变I区)主题和RFLP(限制性片段长度多态性)信息在表2中给出。在这个遗传主题的基础上,我们分析了mtDNA单倍群B4*、B4a、B5a、N9a、F*、F1a、R9b和M7(图5)。mtDNA的HVS-1主题取自有关文献资料。mtDNA的网络全都呈现“太阳”形,由巨大的核心单倍型和四周围绕的大量“日光”般的单倍型所组成。可以看到各语言群体之间错综复杂的融合关系。在F*、M7和N9a的网络中,平话的样本与其他汉族群体关系最密切,表明平话人保持着比汉族父系成分更多的源自汉族母系的成分。但是就总体而言,平话人群的母系遗传仍然与南方少数民族更为接近。
混合分析
汉族起源于华北(山西省一带)并在2500年前开始向南方地区扩展。因而南方汉族遗传上可能的来源是北方汉族和南方少数民族(主要是壮侗和苗瑶族群)。华南汉族群体中这两种亲本族群的混合比率可以用混合分析进行评估。我们使用两种程序(ADMIX2.0和LEADMIX)来估算平话人群父系和母系遗传谱系的混合比。我们假定混血始于2500年前,鉴于其他汉族群体在遗传上相互都很类似,我们将那些样本(1693个Y染色体和2159个mtDNA个体,见表3)汇总为一个可能的亲本人群。我们汇总了华南全部的壮侗和苗瑶族群作为另一可能的南方少数民族亲本族群,(1677个Y染色体和2374个mtDNA个体)。表3清楚地显示出两种程序得出了基本相同的结果。无论在父系还是母系遗传上,南方少数民族对平话人群的贡献率都大大高于汉族群体,表3中所示汉族比率为负值,也许意味着汉族对南方少数民族的贡献率甚至高于对平话人群的贡献率。STR网络所反映的平话人群与两个亲本族群的关系同样也包含在表3中。在Y染色体网络中,平话样本与南方少数民族共享或邻接单倍型明显很多,超过了与汉族共享或邻接的数量。而在mtDNA网络中,平话人群却不特别接近南方少数民族。作为结论,我们主张平话人群的父系主要来自南方少数民族,而母系受到其他汉族群体的影响。
讨论(节选)
……
然而我们未能证实现代平话人群继承了古代平话人群的遗传结构。平话人群可能起源于早期汉族移民,并且保持着源于汉族的核心遗传成分。例如在O3a5a(汉族中常见的单倍群)的网络中(图4),平话样本更接近其他汉族样本而不是南方少数民族,而且有些平话样本处于网络的末端。这意味着平话人群与其他汉族群体分离后,两者的父系遗传谱系各自独立地发展了若干世代。因此也存在另一种可能性:古汉人(从遗传和文化上定义)在广西定居下来之后,曾发生过来自周边原住族群的大规模基因流动,从而使平话人群的古汉族遗传特征显著淡化。也就是说,平话人可能是“遗传上转变了的”汉族移民后裔。
看这种学术论文头都大了
直接告诉贝侬们,咱们的祖先是谁就行了,哈哈
其实我最感兴趣的是华南汉族的演变过程,哪位有才贝侬能说一说?

有人研究过非洲与澳洲人的基因,请将二者联系来做点工作吧。
人类的起源:非洲->印度->印尼->越南->广西->中原
|
->印尼岛屿->澳大利亚
东非过来的都是?那柳江人(柳江的全部人骨化石可能属于同一个中年男性个体。生物分类上归于晚期智人。柳江人是中国以至整个东亚迄今所发现的最早的晚期智人。一般认为其生存年代在距今3—5万年间。)是不是年代上有差别???有矛盾~~关于是否是东非过来的可以怀疑~~~
这是我看完后的总结他写的简短下来后:
所有台湾样本的总体年代估值为21268?148年。有趣的是,后面这个年代值很接近台湾已发现的最早的人类遗存,即左镇人。由此我们推断O1a*个体从大陆向台湾岛的移民出现在旧石器时代.
O1a*单倍群的网络图解释了印尼人的世系并非如语言学研究结果的那样源自台湾原住民,而是来自壮侗族群。
另外,我们的还发现台湾原住民和印尼人在父系上很可能都源自壮侗族群。
相反地,没有台湾世系(除了一个)分享或者连接到东南亚世系的节点。这些发现表明东南亚岛民并非直接源自台湾原住民,而应该是东南亚岛民和台湾原住民各自独立地起源于壮侗群体。
以下这个跟《壮泰传统文化对比研究》有冲突的。
在O2a*的网络中,黎族的分支比大陆和台湾的部分更大,说明黎族是比大陆壮侗族群和台湾原住民更古老,而不是源自大陆壮侗族系的群体。
海南岛上的O2a*分支的年代已确定约为2.6万年前,这个数据比海南最古老的三亚落笔洞遗址更久远。我们提出这就是海南原住民被隔离的年代。
以下这个跟《壮泰传统文化对比研究》有冲突的。
在O2a*的网络中,黎族的分支比大陆和台湾的部分更大,说明黎族是比大陆壮侗族群和台湾原住民更古老,而不是源自大陆壮侗族系的群体。
海南岛上的O2a*分支的年代已确定约为2.6万年前,这个数据比海南最古老的三亚落笔洞遗址更久远。我们提出这就是海南原住民被隔离的年代。
这段话的意思是不是可以理解成这样:海南岛的黎族分支先到达了海南岛,然后大陆和台湾的侗台族群才来到北部湾一带?
我认为可以这样理解:
1.海南岛的新亚洲人移民首先是2万多年前从西面北部湾一带到达的O2a人群。他们在今天的直系后代大概是苗瑶民族和尼科巴岛的匈蓬人,以Y-O2a和mt-B5a为特征标记。
2.早期的O1可能在桂东和粤西一带产生(但是否由壮族的O*中产生还不太确定)。在他们继续向东移民的过程中产生出O1a,从原文看产生地点不会超出从粤东至台湾之间这个范围(包括大陆架)。
3.有一部分O1a群体向西回流,抵达现在的海南岛及其周边地区。这时的O1a可能还不是百越,称之为原始马来人更为合适。
4.进入新石器时代后,海南原住民和现今其他西部的百越民族一样,被西扩的东部百越同化(主要是文化同化),成为现在的黎族。
东非过来的都是?那柳江人(柳江的全部人骨化石可能属于同一个中年男性个体。生物分类上归于晚期智人。柳江人是中国以至整个东亚迄今所发现的最早的晚期智人。一般认为其生存年代在距今3—5万年间。)是不是年代上有差别???有矛盾~~关于是否是东非过来的可以怀疑~~~
不用怀疑人类的非洲起源,因为非洲最早出现的现代人有12-16万年的历史。
这是我看完后的总结他写的简短下来后:
所有台湾样本的总体年代估值为21268?148年。有趣的是,后面这个年代值很接近台湾已发现的最早的人类遗存,即左镇人。由此我们推断O1a*个体从大陆向台湾岛的移民出现在旧石器时代.
O1a*单倍群的网络图解释了印尼人的世系并非如语言学研究结果的那样源自台湾原住民,而是来自壮侗族群。
这里的“壮侗族群”原文中写成Daic,应该不是指现代的壮侗民族,而是指壮侗族群中含有古老的O1a成分。我认为其意思并不确切,因为那时既没有壮侗民族,也没有百越民族,而只有他们的祖先。也许理解成百越、台湾、南岛共同的祖先人群更为合适(我更愿意把他们称为原始马来人)。要知道现在的壮侗傣民族都不是以O1为主体标记的民族。
另外,我们的还发现台湾原住民和印尼人在父系上很可能都源自壮侗族群。
相反地,没有台湾世系(除了一个)分享或者连接到东南亚世系的节点。这些发现表明东南亚岛民并非直接源自台湾原住民,而应该是东南亚岛民和台湾原住民各自独立地起源于壮侗群体。
这句话仍然是就文中所分析的O1a而言,不涉及O2a。
以下这个跟《壮泰传统文化对比研究》有冲突的。
在O2a*的网络中,黎族的分支比大陆和台湾的部分更大,说明黎族是比大陆壮侗族群和台湾原住民更古老,而不是源自大陆壮侗族系的群体。
海南岛上的O2a*分支的年代已确定约为2.6万年前,这个数据比海南最古老的三亚落笔洞遗址更久远。我们提出这就是海南原住民被隔离的年代。
以上两段是李辉《海南原住民》论文中的话,而且全都是就黎族的O2a成分而言的。意思是说:海南的O2a历史远比壮侗O2a的历史久远,因此另有起源。
而《南岛和壮侗》一文主要谈O1a。虽然O1a的整体历史有2万多年,但是海南原住民的O1a只有1万多年。因此海南O1a更像是直接来自大陆上的壮侗族群。这里的壮侗族群(Daic)实际还是在指代百越、台湾、南岛的祖先人群。
以下这个跟《壮泰传统文化对比研究》有冲突的。
在O2a*的网络中,黎族的分支比大陆和台湾的部分更大,说明黎族是比大陆壮侗族群和台湾原住民更古老,而不是源自大陆壮侗族系的群体。
海南岛上的O2a*分支的年代已确定约为2.6万年前,这个数据比海南最古老的三亚落笔洞遗址更久远。我们提出这就是海南原住民被隔离的年代。
这段话的意思是不是可以理解成这样:海南岛的黎族分支先到达了海南岛,然后大陆和台湾的侗台族群才来到北部湾一带?
回:你讲得对。他的意思是黎族要比壮侗群系起源更早。而且是首先到海岛后被隔离(隔离就是不向迁徙的那批)。然后-南群岛和台湾原住民都源自大陆壮侗族系的群体。但黎族却被认为比壮侗族系的群体更古老(的观点)。
关于baiyueren的:这里的“壮侗族群”原文中写成Daic,应该不是指现代的壮侗民族,而是指壮侗族群中含有古老的O1a成分。我认为其意思并不确切,因为那时既没有壮侗民族,也没有百越民族,而只有他们的祖先。
这个李辉他应该最清楚是不是壮侗族。应该至少包含了壮侗族。而Daic正好很可能是壮傣语南部或下方的意思/李辉他承认自己是跟傣族有语言上的相同点,还有民俗等等。而壮侗系群起源于北部湾一带,那正是广西的地方。他没有说是壮族和侗族但他只能用“壮侗系群”代替了。
直到李辉到云南傣族去采样时才恍然发现,傣族的语言他几乎都能听懂;而与水族人交流,李辉更为惊讶,连那儿的悄悄话、私房话,他都能听懂。回到上海后,他就给自己抽血分析,发现傣族、水族和他自己身上都带有M119突变。具有M119突变的族群在民族学上又称为百越民族系统,他们历史上的覆盖范围是从越南北部的交趾分布到江浙一带。在千百年的分化中,诞生了黎族、侗族、水族、仫佬族、仡佬族、高山族、壮族、傣族。这些民族在语言上、文化上有很强的一致性,所以也统称越人。“你看我的眉骨向下弯曲的弧度很大,这就是澳泰语系人群的显著特征。”李辉说。
怀疑造就一切,有怀疑创造智慧~~如果都承认起源于非洲估计要等很多人多年的证实~因为他里面写的用了很多“可能”!
不过这些资料是丰富了很多人的思维~~
其实李辉的意思已经很清楚了。在描述已经混血消失的古代族群时,任何人都会感到语汇的贫乏和不确切。
说得再清楚一点,现代的百越族群至少有两种基本成分:一种是古代的O1*人群,他们的直系后代是几个台湾部族(如泰雅族),浙江的O1比例也可以达到约25%。另一种是古代的O2a人群,他们的直系后代是现代的瑶族和尼科巴岛匈蓬人。东部百越的O1成分较多,而西部百越的O2a成分更多。
从现有论文资料看,百越民族很可能形成于广东一带,也就是已经消失的东部百越才是真正的源头。现在的黎、壮、侗、傣,包括台湾原住民族,其实都是古代百越同化和兼并其他族群的结果。其中最主要同化对象就是O2a族群。所以我认为应该把现代壮侗傣族群看成是百越同化的结果(而不是源头)更为准确。
当然,还有一个重要的问题没有解决,就是壮族的O*所代表的古代族群是不是O1、O2a和O3的祖先?如果整个发展的脉络都清楚了,才更有把握地回答百越-原始马来族群的起源问题。
现在的研究结果,似乎是O*更古老一些,呵呵。还有一个问题,可能在5000甚至是3000年以前,百越族群很有可能仍旧是母系氏族社会,因些,不能光以Y染色体来断定他们的遗传关系,还须研究母系基因的遗传情况,期待这方面的研究结果。
原来,你们真是马来人,马来猪。
你们和马来猴子是同胞啊!难怪老是拿汉人说事。
楼上的“欧阳子燕”网友已经涉嫌违规,请请管理员记录在案。
| 欢迎光临 僚人家园 (http://bbs.rauz.net.cn/) | Powered by Discuz! X2.5 |